Written by丨Enzyme Beauty
The mTORC1 complex is the switch for cellular anabolic metabolism, sensing and integrating the effects of cellular energy, nutrient levels, and growth factor stimulation to promote cell growth while inhibiting catabolism. Under nutrient-rich conditions, the Ragulator-Rag GTPase is responsible for recruiting mTORC1 to the surface of lysosomes, the organelles that execute the function of biomolecule degradation. The cellular nutritional state regulates mTORC1 activity through Rag GTPases. Changes in amino acid levels are sensed by receptors-GATOR2-GATOR1, which control the GTP-GDP switching of Rag GTPase RagA/RagB. Meanwhile, FLCN-FNIP2 and leucyl-tRNA synthetase regulate RagC/RagD and RagD.
When active, Rag GTPases form a dimer, and must be a heterodimer of RagA/B (GTP):RagC/D (GDP) to recruit mTORC1. RagA/B and RagC/D have high homology in their main functional domains and have been considered functionally interchangeable. However, they differ significantly in their N-terminal and C-terminal sequences, providing a basis for differential regulation. Previous studies indicated that leucyl-tRNA synthetase could only regulate RagD, further suggesting possible differences among the four Rag proteins.
Recently, a team from the Max Planck Institute for Biology of Ageing in Germany, led by Constantinos Demetriades, and a team from the Cologne Center for Genomics, led by Aurelio A. Teleman, published two back-to-back studies in Nature Cell Biol, titled:A Rag GTPase dimer code defines the regulation of mTORC1 by amino acids and Brain-enriched RagB isoforms regulate the dynamics of mTORC1 activity through GATOR1 inhibition. Both studies demonstrate the comprehensive differences of Rag: different affinities for the mTORC1 complex, varied responses to nutritional states, distinct distributions across tissues leading to organ differentiation, and differential regulation of downstream substrates. Through different Rag combinations, mammals achieve spatiotemporal differential regulation of mTORC1 in various scenarios.
Both teams utilized Rag knockout cell lines and expressed different combinations of Rag dimer genes to explore the differences among Rags.
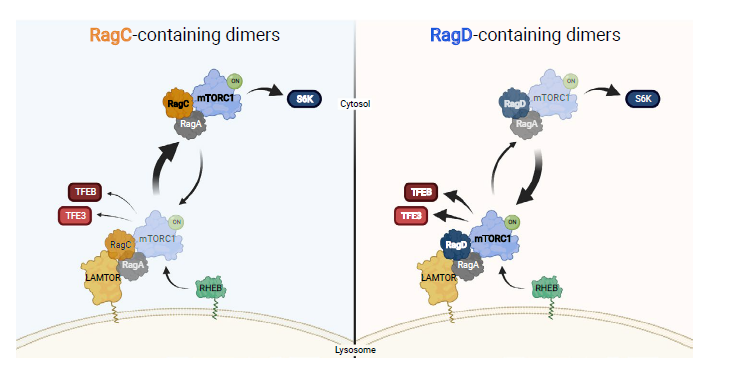
The Demetriades team constructed a Rag knockout 293FT cell line and then established stable Rag-expressing cells. They observed that RagA/D expression can restore mTORC1 lysosomal localization and phosphorylation of downstream substrate TFEB/TFE3, while TFEB/TFE nuclear localization disappears. In the RagA/C combination, there were fewer mTORC1 lysosomal localization signals; although the phosphorylation level of downstream substrate S6K significantly increased, RagA/C could hardly detect TFEB/TFE3 phosphorylation signals, yet some cells still showed TFEB/TFE3 nuclear entry. As TFEB/TFE3 nuclear entry decreased, corresponding lysosome biogenesis and hydrolase production also decreased. Consistent with this, in the RagA/D combination, lysosome biogenesis and hydrolase production returned to wild-type cell levels, while in RagA/C combination cells, a certain level of lysosomes and lysosomal hydrolase increase was still observed.
The Demetriades team further traced the differential regulatory abilities of RagC and RagD on TFEB/TFE3. Co-immunoprecipitation experiments detected that the RagD subunit had a stronger binding affinity to p18/LAMTOR1, and RagD heterodimers were more recruited to lysosomes. Using Rag subunits as bait, lysosomal precipitation LysoIP indicated that RagD precipitation would acquire more lysosomes. Sequence swapping experiments demonstrated that the N-terminal and C-terminal non-structured domain sequences of the RagD subunit conferred a stronger binding affinity to LAMTOR. The study also found that cancer mutations in RagC, RagCT90N and RagCW115R acquired superpowers similar to RagD, increasing mTORC1 activity and TFEB/TFE3 phosphorylation, corroborating the RagD experimental results.
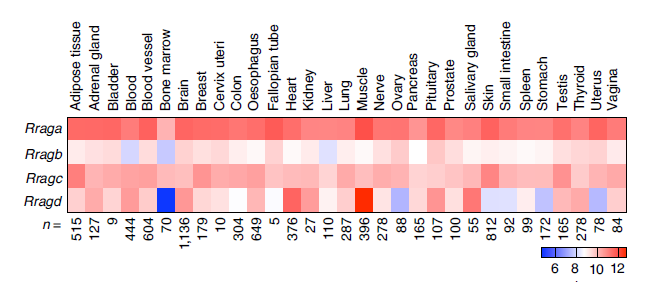
 The Teleman team first observed the different distributions of the four Rag proteins across tissues, with RagB having both long and short isoforms, where RagBshort is highly expressed in brain tissue. It has long been observed that under nutrient-deficient conditions, fetal development in utero shows some stagnation while brain development remains unaffected. A similar phenomenon occurs under hypoxic conditions in the fetal brain. Therefore, the team explored the RagB subunit.
The Teleman team first observed the different distributions of the four Rag proteins across tissues, with RagB having both long and short isoforms, where RagBshort is highly expressed in brain tissue. It has long been observed that under nutrient-deficient conditions, fetal development in utero shows some stagnation while brain development remains unaffected. A similar phenomenon occurs under hypoxic conditions in the fetal brain. Therefore, the team explored the RagB subunit.
 The team constructed a RagA/B double knockout cell line and stably expressed either RagA or RagB genes. Under amino acid scarcity, Rag A/B switched to GDP binding, losing mTORC1 lysosomal localization and reducing downstream substrate phosphorylation. The team observed significant differences in the responses of cells expressing RagA and RagB to changes in amino acid levels. Under amino acid starvation conditions, cells expressing RagBshort and RagBlong retained mTORC1 localization at lysosomes, with downstream substrates S6K, TFEB/TFE3, and 4EBP-1 phosphorylation levels largely maintained, indicating that RagB can resist the suppression of Rag GTPase under nutrient deficiency, maintaining partial mTORC1 activity and ensuring cellular biosynthesis. RagBshort and RagBlong exhibited different performances under nutrient-rich conditions. RagBlong showed insufficient recruitment to mTORC1 and weaker binding to Raptor.
The Teleman team next explored the differences in regulatory mechanisms between RagA/B. Changes in amino acid levels control the activity of RagA/RagB through receptors-GATOR2-GATOR1, and the study found that GATOR1 exerted weaker control over the two isoforms of RagB. Results from sequence swapping and in vitro GAP activity experiments indicated that RagBshort had more binding to the inhibitory domain of the GATOR1 component DEPDC5, thus inhibiting GATOR1’s GAP activity. RagBlong primarily formed strong associations with GATOR1 in a non-GTP binding state, functioning similarly to a sponge, sequestering GATOR1 away from RagA/B proteins. At the same time, RagBlong could inhibit GATOR1’s GAP activity, and there was a synergistic effect in the inhibitory actions of RagB’s two isoforms on GAP activity.
In summary, the Teleman team’s exploration of RagB illustrates that due to RagB’s resistance to GATOR1 activity suppression, and its high expression in brain tissue, our brains maintain a certain level of mTORC1 activity under stress conditions, allowing biosynthesis to continue in brain tissue. Unfortunately, studies from TGCA and CCLE found that the expression ratio of RagB/RagA is high in certain cancer types, suggesting that tumor tissues can increase RagB expression to ensure biosynthesis under nutrient-deficient growth conditions.
As studies published back-to-back, both teams also validated each other’s main findings.These two NCB studies present detailed experimental data showcasing the fine spatiotemporal regulation of mTORC1 activity by biology, with RagA/B-RagC/D forming a biphasic logic gate for mTORC1 activity regulation, achieving fine-tuned control of mTORC1 activity through differences in Rag expression, distribution, and protein interactions.
The team constructed a RagA/B double knockout cell line and stably expressed either RagA or RagB genes. Under amino acid scarcity, Rag A/B switched to GDP binding, losing mTORC1 lysosomal localization and reducing downstream substrate phosphorylation. The team observed significant differences in the responses of cells expressing RagA and RagB to changes in amino acid levels. Under amino acid starvation conditions, cells expressing RagBshort and RagBlong retained mTORC1 localization at lysosomes, with downstream substrates S6K, TFEB/TFE3, and 4EBP-1 phosphorylation levels largely maintained, indicating that RagB can resist the suppression of Rag GTPase under nutrient deficiency, maintaining partial mTORC1 activity and ensuring cellular biosynthesis. RagBshort and RagBlong exhibited different performances under nutrient-rich conditions. RagBlong showed insufficient recruitment to mTORC1 and weaker binding to Raptor.
The Teleman team next explored the differences in regulatory mechanisms between RagA/B. Changes in amino acid levels control the activity of RagA/RagB through receptors-GATOR2-GATOR1, and the study found that GATOR1 exerted weaker control over the two isoforms of RagB. Results from sequence swapping and in vitro GAP activity experiments indicated that RagBshort had more binding to the inhibitory domain of the GATOR1 component DEPDC5, thus inhibiting GATOR1’s GAP activity. RagBlong primarily formed strong associations with GATOR1 in a non-GTP binding state, functioning similarly to a sponge, sequestering GATOR1 away from RagA/B proteins. At the same time, RagBlong could inhibit GATOR1’s GAP activity, and there was a synergistic effect in the inhibitory actions of RagB’s two isoforms on GAP activity.
In summary, the Teleman team’s exploration of RagB illustrates that due to RagB’s resistance to GATOR1 activity suppression, and its high expression in brain tissue, our brains maintain a certain level of mTORC1 activity under stress conditions, allowing biosynthesis to continue in brain tissue. Unfortunately, studies from TGCA and CCLE found that the expression ratio of RagB/RagA is high in certain cancer types, suggesting that tumor tissues can increase RagB expression to ensure biosynthesis under nutrient-deficient growth conditions.
As studies published back-to-back, both teams also validated each other’s main findings.These two NCB studies present detailed experimental data showcasing the fine spatiotemporal regulation of mTORC1 activity by biology, with RagA/B-RagC/D forming a biphasic logic gate for mTORC1 activity regulation, achieving fine-tuned control of mTORC1 activity through differences in Rag expression, distribution, and protein interactions.
Editor: Eleven
1. https://doi.org/10.1038/s41556-022-00976-y
2. https://doi.org/10.1038/s41556-022-00977-x
【Original Article】BioArt Original Article, individual sharing is welcome, but reproduction without permission is prohibited. All published works are owned by BioArt. BioArt reserves all legal rights, and violators will be prosecuted.








