From clicking the blue text above ↑↑↑——“Stem Cell Talk“, mark with a star to follow this popular science account.
Neither praise nor blame
To state the facts
Is the way of popular science
There is still a long way to go from basic research to clinical application, but this is a process that every new technology must undergo, and basic research is essential.
Written by: Picky Immunity Cat
Source: Casual Talk Immunology

1
The evolution of species diversity and survival struggles has produced a complex host defense system that relies on innate immunity, first in unicellular eukaryotes, then adaptive immunity emerging in multicellular and developing eukaryotes. The concept of innate immunity was first articulated at the end of the 19th century when Elie Metchnikoff introduced the term “macrophage,” meaning “macro = big and phage = eater“. Monocytes/macrophages are a small group of leukocytes defined by their location, phenotype, morphology, and gene expression profile.
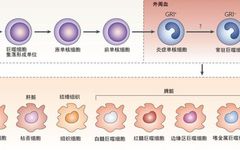
Monocytes account for 4-10% of nucleated cells in normal human peripheral blood. In the bloodstream, the half-life of monocytes is less than 20 hours. For many years, it was assumed that macrophages were entirely derived from circulating monocytes, but recent morphological and functional differences between these cells have refuted this hypothesis.
Recent research evidence suggests that most macrophages in adult tissues are seeded prenatally, extracted from the yolk sac during embryonic development, possess self-renewal capabilities, and are maintained independently of monocytes. Furthermore, each organ has its unique combination of embryonic and adult-derived macrophages.
Embryonic macrophages participate in tissue remodeling, while adult macrophages primarily assist in host defense. Aside from these differences, we also observe the coexistence of embryonic and adult macrophages in many different organs.
2
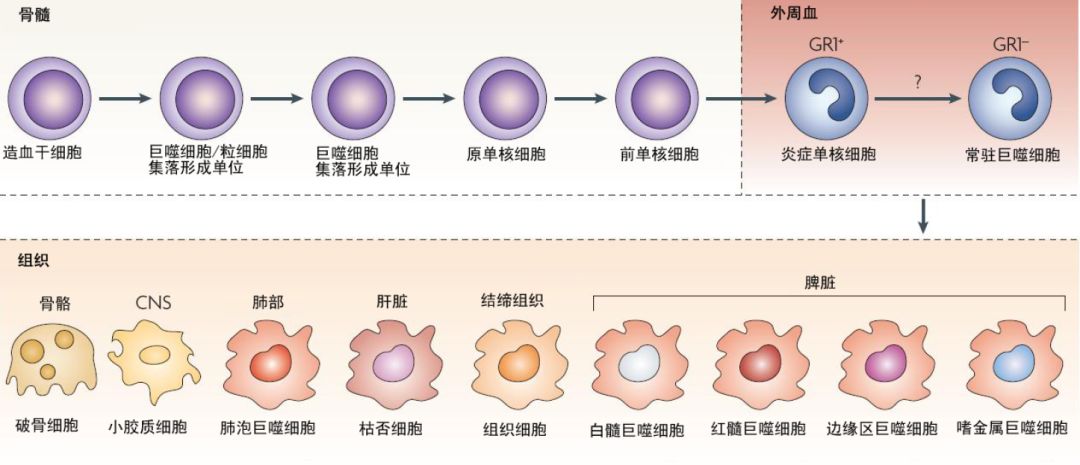
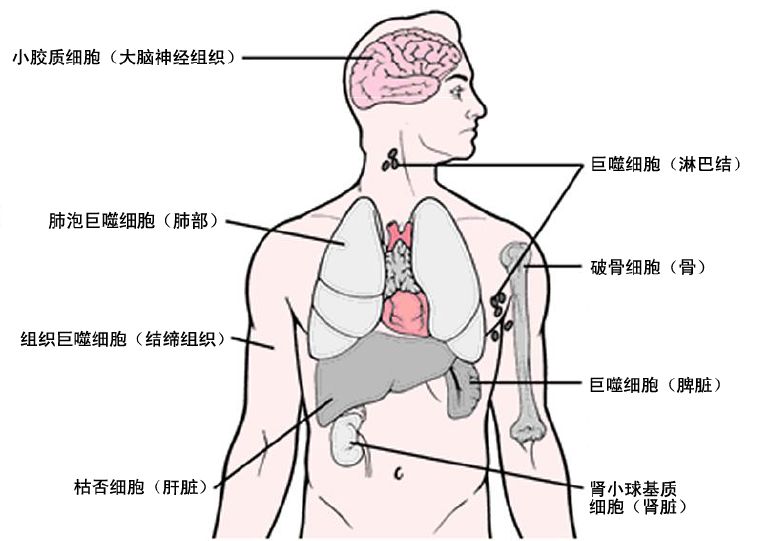
Macrophages are classified based on their anatomical location and functional phenotype into microglia, osteoclasts, pulmonary macrophages, splenic tissue macrophages, mesenchymal connective tissue, liver mesenchymal connective tissue, and Kupffer cells.
There are also different types of macrophages in the intestines, which exhibit different phenotypes and functions but work together to maintain tolerance to the normal gut microbiota and oral antigens.
Secondary lymphoid organs also contain varying numbers of macrophages, including marginal zone macrophages in the spleen that suppress natural and adaptive immune responses to apoptotic cells, and subcapsular sinus macrophages in lymph nodes that clear viruses from lymph and initiate antiviral immune responses.
Different macrophages exist in immune privileged sites, such as the brain, eyes, and testes, which play a central role in tissue remodeling and dynamic balance. These tissue-specific macrophages engulf dead cells, debris, foreign antigens, and materials, organize the inflammatory process, and recruit more macrophages as needed.
3
Macrophages are highly plastic cells that can switch from one phenotype to another. Macrophage polarization is a process where macrophages exhibit specific phenotypes and respond functionally to the stimuli and signals encountered in each specific tissue microenvironment.
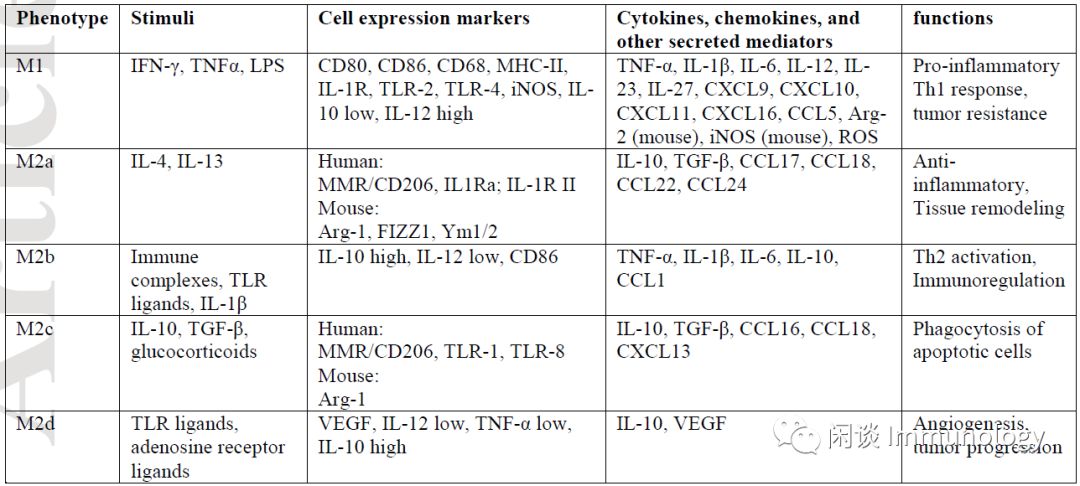
The local cytokine environment can dictate macrophage polarization. The two major macrophage subpopulations with different functions include classically activated or inflammatory (M1) and alternatively activated or anti-inflammatory (M2) macrophages. This phenomenon of the two different M1/M2 phenotypes is termed macrophage polarization.
M1 macrophages are typically induced by Th1 cytokines (such as IFN-γ and TNF-α) or bacterial lipopolysaccharides (LPS). These macrophages produce and secrete higher levels of pro-inflammatory cytokines TNF-α, IL-1α, IL-1β, IL-6, IL-12, IL-23, and cyclooxygenase-2 (COX-2), while having lower levels of IL-10.
Functionally, M1 macrophages participate in the clearance of pathogens during infection by activating the nicotinamide adenine dinucleotide phosphate (NADPH) oxidase system and subsequently generating reactive oxygen species (ROS). Therefore, M1 macrophages exhibit strong antimicrobial and antitumor activity, mediating tissue damage caused by reactive oxygen, impairing tissue regeneration and wound healing. To prevent such tissue damage, the anti-inflammatory effects of M2 macrophages suppress chronic inflammatory responses through regulatory mechanisms.
M2 macrophages are activated by IL-4 receptor α (IL-4Rα), polarized by Th2 cytokines IL-4 and IL-13, and exhibit anti-inflammatory effects. In addition to IL-4 and IL-13, cytokines such as IL-10 can also regulate M2 polarization by activating STAT 3 through the IL-10 receptor (IL-10R).
IL-33 is a cytokine associated with Th2-related cytokines in the IL-1 family that can induce M2 polarization. It is characterized by the upregulation of arginase-1 (arg-1), CCL 17, and CCL 24, mediating eosinophilia and airway inflammation.
IL-21 is another Th2-related cytokine that drives M2 polarization. M2 macrophages are characterized by the production of anti-inflammatory cytokines, characterized by low IL-12 production and high levels of IL-10 and TGF-β.
Functionally, M2 macrophages exhibit strong phagocytic abilities, clearing debris and apoptotic cells, promoting tissue repair and wound healing, and exhibiting angiogenic and fibrotic properties. Therefore, in general, M2 cells participate in Th2 responses and parasite clearance, suppress inflammation, promote tissue remodeling, angiogenesis, immune regulation, tumor formation, and progression.
However, the M1/M2 phenotypes do not reflect the different phenotypic subgroups of macrophages. Depending on the activating stimuli received, M2 macrophages can be further divided into four different subgroups composed of M2A, M2B, M2C, and M2D.
M2a macrophage subgroup can be induced by IL-4 and IL-13, producing high levels of CD206, decoy receptor IL-1 receptor II (IL-RII), and IL-1 receptor antagonist (IL-1Ra).
Immune complexes (ICs) and Toll-like receptor (TLR) agonists or IL-1 receptor ligands can induce the M2b subgroup. M2b macrophages produce both anti-inflammatory and pro-inflammatory cytokines IL-10, IL-1β, IL-6, and α-α.
M2c cells are induced by glucocorticoids and IL-10, exhibiting strong anti-inflammatory effects on apoptotic cells by releasing large amounts of IL-10 and transforming growth factor-β.
Finally, TLR agonists induce the fourth type of M2 macrophage M2d through adenosine receptor agonists.
The activation of adenosine receptors subsequently inhibits the production of pro-inflammatory cytokines, inducing the secretion of anti-inflammatory cytokines (high IL-10, low IL-12) and vascular endothelial growth factor (VEGF), thus providing vascularization characteristics typical of tumor-associated macrophages (TAMs). The exposure of M2 macrophages to M1 signals, or vice versa, induces differentiated macrophages to “repolarize” or “reprogram,” providing further evidence of their high functional plasticity, which can potentially pursue therapeutic targets.
4
Macrophage Migration
Monocytes and macrophages migrate to sites of inflammation or injury to eliminate the initial inflammatory signals, ultimately promoting wound healing and tissue repair.
This process is primarily initiated by pathogen-associated molecular patterns (PAMPs) (released from invading pathogens) and damage-associated molecular patterns (DAMPs) (released from damaged or dead cells in response to infection and injury).
Moreover, antigen-activated tissue-resident memory T cells can trigger macrophage recruitment by secreting multiple inflammatory cytokines and chemokines. Chemokines directly participate in the migration and activation of monocytes through the endothelium, with monocyte chemotactic protein-1 (MCP-1) being a monocyte chemokine involved in the occurrence of inflammation, inducing monocyte chemotaxis and migration by interacting with CC chemokine receptor 2 (CCR2) on monocytes. MCP-1 is primarily secreted by activated fibroblasts, endothelial cells, vascular smooth muscle cells (VSMC), monocytes, and T cells. Together with IL-8 or CXC ligand-8 (CXCL-8), MCP-1 can trigger the firm adhesion of monocytes to vascular endothelial cells under flow conditions. The adhesion of monocytes and transendothelial migration through activated venular walls is a fundamental immune response that relies on the expression of adhesion molecules by chemical mediators on the venous endothelium.
These adhesion molecules belong to four families: selectins, integrins, immunoglobulin superfamily, and mucin-like glycoproteins.
5
Antigen Presentation
Antigen-presenting cells, primarily macrophages, are the sentinels of the immune system that initiate and regulate immune responses. An important feature of macrophage biology is the capture, uptake, and expression of self or foreign antigens, which provides the link between innate and adaptive immunity.
Macrophages reside in surrounding organs, where they monitor invading pathogens in surrounding tissues. They engulf pathogens, process their antigens, and present peptide fragments bound to human leukocyte antigen (HLA) molecules, alerting the immune system to the presence of pathogens. After antigen processing, macrophages migrate to T cells and stimulate them. Activated macrophages express high levels of co-stimulatory molecules and antigen-presenting molecules on their surface, such as CD 80, CD 86, and MHC class I and II molecules. Mixed leukocyte/lymphocyte reactions (MLR) are used as a basic test for assessing macrophage function, as they measure the proliferation capacity of macrophages against allogeneic T cell populations.
6
Cytokine and Chemokine Production
Interleukins are a class of cytokines involved in inducing adhesion molecules, matrix metalloproteinases (MMPs), pro-angiogenic factors, and signaling pathways, such as nuclear factor kappa B (NF-kB) and signal transducer and activator of transcription (STAT) involved in tumor invasion and angiogenesis.
M1 macrophage activation can induce pro-inflammatory cytokines, including TNF-α, IL-1α, IL-1β, IL-6, IL-12, IL-18, and IL-23; produce nitric oxide (NO), reactive oxygen species (ROS), and RNS; antigen presentation; and express CC chemokine receptors CCR1 and CCR5; promoting T helper type 1 (Th1) and Th17 responses, providing an effective mechanism for killing.
Clearly, M1 and M2 macrophages have different chemokine profiles, with M1 macrophages expressing Th1 cell chemokines such as CXCL9 and CXCL10.
M2 macrophages express chemokines CCL17, CCL18, CCL22, and CCL 24. M1 macrophages express inflammatory cytokines TNF-α, IL-1β, IL-6, and upregulate MHC class II molecules together with co-stimulatory molecules CD 40, CD 80, and CD 86, promoting cytotoxic adaptive immunity.
M1 macrophages express Th1- and Th17-polarizing cytokines IL-12, IL-23, IL-27, and Th1-attracting chemokines CXCL 9, CXCL 10, CXCL 11. In contrast, M2 macrophages support the resolution of inflammation by shifting gene expression toward anti-inflammatory molecules such as IL-10, TGF-β, IL-1R type II, and IL-1Ra. M2 macrophages also express a large number of endogenous receptors, including scavenger receptor CD 163, stabilin-1, and C-type lectin receptors CD 206, CD 301, Dectin-1, and CD 209. Furthermore, M2 macrophages recruit Th2, regulatory T cells (Tregs), eosinophils, and basophils by secreting CCL 17, CCL 18, CCL 22, and CCL 24 chemokines.
7
Macrophage Apoptosis
Apoptosis is a normal, physiological, and necessary process of cell selection and survival, eliminating and removing unwanted cells, such as senescent, damaged, genetically mutated, and virus-infected cells; in fact, anti-apoptosis can promote the malignant transformation of normal cells.
It is now known that macrophage apoptosis is associated with various stages of diseases related to the decline in pathogen viability, which can be seen as a natural immune defense mechanism and strategy used by the host to limit infections.
Conversely, many intracellular pathogens have the ability to kill infected cells, disrupt host pathways, and inhibit apoptosis, leading to necrosis. This strategy allows pathogens to evade host immunity and infect other nearby or distant cells.
These apoptotic infected cells also provide a connection between innate and adaptive immunity. This connection begins when professional APCs absorb apoptotic vesicles, enhancing T cell activation through bacterial products and pathogenic antigens. Similarly, pathogens that inhibit apoptosis and promote necrosis can impair the presentation of bacterial antigens, which may be mechanisms of immune evasion.
In atherosclerosis, macrophages undergo apoptosis by reducing the number of cells within lesions, favoring a reduction in lesion size. During the progression of atherosclerosis and other diseases, the phagocytic function of M2 macrophages resolves apoptosis and its adverse effects. This unique functional characteristic of M2 macrophages is supported by the high expression of scavenger receptors.
Macrophages engulf debris, damaged cells, dead cells, and apoptotic cells, inhibiting the production of pro-inflammatory cytokines such as tumor necrosis factor-α, interleukin-1β, interleukin-6, and interleukin-8, through mechanisms involving the autocrine or paracrine secretion of transforming growth factor-β (TGF-β), thereby inhibiting further recruitment of monocytes and macrophages.
Moreover, the engulfment of apoptotic cells suppresses the production of pro-inflammatory cytokines IL-12, IL-23, and IL-27 and stimulates the production of the anti-inflammatory cytokine IL-10.
By limiting the phagocytic action of local inflammation, it can also protect tissues from harmful pro-inflammatory responses, death, damage, and the immunogenic content of dead and dying cells, as well as protect lesions from growth through necrosis after apoptosis.
In the late stages of atherosclerosis, the phagocytic function of apoptotic cells is severely impaired, leading to the accumulation of macrophages and vascular smooth muscle cells, ultimately blocking the resolution of inflammation, promoting local necrosis, inflammatory states, and plaque instability.
Understanding the exact mechanisms by which pathogens suppress apoptosis to evade innate and adaptive immunity offers new strategies for optimizing therapies. The idea of manipulating host or bacterial pathways to induce more apoptosis, termed “apoptosis-promoting therapy,” may help generate more effective immune responses.
8
Macrophage Metabolism
Recent evidence suggests that metabolism plays an important role in shaping macrophages’ apparent polarized phenotypes. Under normal, physiological, and pathological conditions, macrophages face oxygen gradients.Macrophages adapt to hypoxia by shifting their metabolic pathways towards glycolysis. Additionally, the activation of hypoxia-inducible factors (HIF)-1 and -2 leads to profound functional changes, including the expression of chemokines and chemokine receptors, such as CXC chemokine 4 (CXCR 4), CXCL 12, angiogenic factors, and vascular endothelial growth factor (VEGF) (Biswas and Mantovani, 2012; Orihuela et al., 2016). Thus, macrophages contribute to coordinating tissue responses to oxygen gradients and hypoxic conditions.
M1 and M2 macrophages exhibit significant regulatory effects on the metabolism of glucose, amino acids, iron, and folate.After macrophage activation, their metabolic pathways shift from oxidative to glycolytic. Under M1 stimulation (such as LPS, Th1 cytokines, and IL-12), they demonstrate a metabolic transition toward anaerobic glycolytic pathways, while M2 stimulation (such as IL-4 and IL-13) has less effect. Macrophages activated by IFN-γ and LPS are often associated with acute infections; therefore, they need to rapidly trigger strong antimicrobial activity in hypoxic microenvironments. In this case, anaerobic processes such as glycolysis are the best pathways when energy is required. Interferon-γ and lipopolysaccharides strongly induce glucose uptake while inhibiting fatty acid uptake and oxidation. Consequently, glycolytic metabolism is enhanced, and mitochondrial activity is reduced.
Conversely, M2 macrophage functions, such as wound healing and tissue repair, require a continuous energy supply. This demand is provided by glucose oxidative metabolism and fatty acid oxidation, while fatty acid oxidation is the most important metabolic pathway for M2 macrophages.
M2 macrophages exhibit higher mitochondrial activity and enhanced oxidative phosphorylation characteristics. They show increased respiratory activity based on fatty acid β-oxidation. Both metabolic pathways are upregulated during macrophage activation, but glycolysis is the primary metabolic pathway that generates substrates for DNA and cell membrane synthesis, promoting monocyte growth and differentiation. Producing rapid and optimal responses at infection sites is another possibility for the glycolytic shift of M1 macrophages.
Furthermore, the accumulation of citrate in M1 macrophages is essential for the production of pro-inflammatory mediators NO, ROS, RNS, and prostaglandins. Another important aspect is the metabolism of amino acids closely associated with macrophage functional phenotypes. M1 macrophages are characterized by the expression of inducible nitric oxide synthase (iNOS).NO production is an important effect of antimicrobial activity in M1 macrophages, while M2 macrophages do not produce NO. In contrast, they express high levels of Arg-1, which plays a role in catalyzing polyamine production, necessary for collagen synthesis, cell proliferation, fibrosis, and other tissue remodeling functions. Interestingly, the production of polyamines has been reported as a driving factor for M2 polarization.
Moreover, there are significant differences in iron metabolism between M1 and M2 macrophages. While M1 macrophages express large amounts of iron storage proteins, such as ferritin, they express lower levels of heme, which is the export form of iron. Compared to M1 macrophages, M2 macrophages have lower ferritin content but higher heme content. This different iron metabolism may relate to their functions. Since iron is essential for supporting bacterial growth, the iron retention in M1 macrophages serves as a bacteriostatic effect and supports host protection against infections. In contrast, the iron released by M2 macrophages favors tissue repair but can lead to tumor growth and metastasis.
Metabolic adaptation is an important aspect of macrophage polarization and their functional activities.
9
MicroRNA Involved in Macrophage Polarization
MicroRNAs play important roles in many aspects of macrophage biology, thereby affecting various biological and pathological conditions such as monocyte differentiation and development, macrophage polarization, infection, atherosclerosis, tumor growth, inflammatory activation, cholesterol homeostasis, cell survival and proliferation, and phagocytosis, etc.
miRNA-24, miRNA-30b, miRNA-142-3p, and miRNA-199a-5p can inhibit the differentiation of monocytes into macrophages.
M1 macrophage polarization requires miRNA-125, miRNA-146, miRNA-155, miRNA-7a/f, and miRNA-378, while M2 polarization requires miRNA-let-7c/e, miRNA-9, miRNA-21, miRNA-146, miRNA-147, miRNA-187, and miRNA-223.
Moreover, miRNA-342-5p promotes NO synthesis, and IL-6 provides pro-inflammatory signals for macrophages. Although miR-21 has been shown to exhibit both pro-inflammatory and anti-inflammatory effects, its anti-inflammatory effect is more pronounced. miR-155 and miR-142-3p inhibit macrophage proliferation compared to let-7a. miR-155 has both pro-apoptotic and anti-apoptotic effects, while miR-21 and let-7e negatively regulate macrophage apoptosis.
Key References
Abbas Shapouri Moghaddam, doi:10.1002/jcp.26429, 2018
Thomas A. Wynn, Nature, doi:10.1038/nature12034, 2013
Nicole J. Horwood, Clinic Rev Allerg Immunol, doi:10.1007/s12016-015-8519-2

Riding the East Wind of Stem Cells – The Rise of “Exosomes”
Related Popular Science:
How to View the Safety Issues of Mesenchymal Stem Cell Therapy?
How to View the Impact of the Microenvironment on Mesenchymal Stem Cell Therapy?
How to View “Stem Cell Therapy for Ovarian Insufficiency”?
How to View “Combination Use of PRP and Stem Cells”?
How to View the Impact of Stem Cell Microenvironment on Regenerative Ability?
Mesenchymal Stem Cell Therapy for Diabetic Foot Ulcers Shows Great Potential
Can Mesenchymal Stem Cells Repair Meniscus Injuries?
Mesenchymal Stem Cells and Graft-versus-Host Disease
Mesenchymal Stem Cells and Human Cartilage Regeneration
Talking About Our Understanding of Stem Cell Therapy Despite the Long Way Ahead
Hair Loss and Baldness: Secrets Hidden in Hair Follicle Stem Cells
The Secret of Human Reproduction: Germline Stem Cells
How to Understand the Homing Ability of Stem Cells Accurately
— END —


Stem Cell Talk
– Popular Science Sentiment Responsibility –