
 Summary:Rabies is an ancient disease that has a history spanning thousands of years since the first interactions between humans and dogs. The astonishing death toll caused by this disease since the first century BC has triggered rabies prevention strategies. Over the past 100 years, countless attempts have been made to develop rabies vaccines to prevent rabies in humans and animals. Pre-Pasteur vaccine scientists paved the way for the practical history of rabies vaccines by developing the first generation of vaccines. Further improvements to low-reactogenic and high-immunogenic vaccines have led to the development of embryonic vaccines, tissue culture vaccines, cell culture vaccines, modified live vaccines, inactivated vaccines, and adjuvant vaccines. The development of recombinant technology and reverse genetics has provided insight into the rabies virus genome and facilitated genomic manipulation, which in turn has led to the emergence of the next generation of rabies vaccines, such as recombinant vaccines, viral vector vaccines, transgenic vaccines, and nucleic acid vaccines. These vaccines are significantly helpful in overcoming the drawbacks of traditional rabies vaccines, enhancing immunogenicity and clinical efficacy. From Pasteur to modern vaccines, the development of rabies vaccines has undergone numerous challenges; this pioneering work has laid the foundation for the successful vaccines currently available for rabies prevention. In the future, advances in science and technology and research focus will undoubtedly pave the way for more complex rabies vaccine candidates.
Summary:Rabies is an ancient disease that has a history spanning thousands of years since the first interactions between humans and dogs. The astonishing death toll caused by this disease since the first century BC has triggered rabies prevention strategies. Over the past 100 years, countless attempts have been made to develop rabies vaccines to prevent rabies in humans and animals. Pre-Pasteur vaccine scientists paved the way for the practical history of rabies vaccines by developing the first generation of vaccines. Further improvements to low-reactogenic and high-immunogenic vaccines have led to the development of embryonic vaccines, tissue culture vaccines, cell culture vaccines, modified live vaccines, inactivated vaccines, and adjuvant vaccines. The development of recombinant technology and reverse genetics has provided insight into the rabies virus genome and facilitated genomic manipulation, which in turn has led to the emergence of the next generation of rabies vaccines, such as recombinant vaccines, viral vector vaccines, transgenic vaccines, and nucleic acid vaccines. These vaccines are significantly helpful in overcoming the drawbacks of traditional rabies vaccines, enhancing immunogenicity and clinical efficacy. From Pasteur to modern vaccines, the development of rabies vaccines has undergone numerous challenges; this pioneering work has laid the foundation for the successful vaccines currently available for rabies prevention. In the future, advances in science and technology and research focus will undoubtedly pave the way for more complex rabies vaccine candidates.
01
Introduction
Since humans and dogs began interacting nearly 40,000 years ago, rabies has held significant historical importance. Records from Mesopotamia reveal the existence of a highly dangerous “mad dog” disease, highlighting the interaction between dogs and the most lethal rabies virus..Rabies is a deadly infectious zoonotic disease caused by the rabies virus..The disease continues to pose a serious threat to global public health, especially in developing countries. This acute progressive encephalitis causes approximately 60,000 deaths annually, primarily in Africa (36.4%) and Asia (59.6%). South Asia accounts for about 40% of the world’s human rabies deaths. It is estimated that rabies causes annual losses of $583.5 million, with livestock losses in Asia and Africa amounting to approximately $12.3 million. Canine rabies is present in 87 different countries and is a major factor in human rabies cases. However, some countries, including Japan, the UK, Denmark, Sweden, Greece, Ireland, Iceland, Portugal, New Zealand, Australia, Switzerland, Finland, Norway, France, and Belgium, have eradicated rabies..
The main permanent transmitter of the disease is the rabies virus (RABV), which is a prototype species in the rabies virus genus of the Rhabdoviridae family. It is a bullet-shaped virus with a single-stranded negative RNA genome of approximately 12 kb, encoding five major structural proteins from 3’ to 5’, namely nucleoprotein (N), phosphoprotein (P), matrix protein (M), glycoprotein (G), and RNA-dependent RNA polymerase (L). The N, P, and L proteins form a ribonucleoprotein complex that tightly encapsulates the negative RNA genome and is responsible for guiding the virus’s replication in the cytoplasm of infected cells. The RABV G protein is the only viral protein exposed on the virus surface and is a major factor leading to the virus’s pathogenicity, serving as a primary protective antigen that induces protective immunity against rabies.
The disease affects all warm-blooded animals, including humans, and the rabies virus has expanded its host range to mammals, including carnivores and bats. See MEGACHIROPTERA. However, among these hosts, dogs are the most important hosts for human infections in developing countries, while wildlife serves as hosts in developed countries..In addition to dogs, several bat species, particularly vampire bats, play a crucial role in the transmission of human rabies virus in the Americas..Conversely, in Old World countries in Africa, Asia, and Europe, the rabies virus is transmitted by bats..Other livestock, including cats, cattle, horses, sheep, and goats, can be infected with rabies and transmit it to humans..Infected dog bites account for 97% of human rabies cases, followed by cat bites (2%) and bites from other animals (1%), including mongoose, foxes, wolves, jackals, and other wildlife..
Fortunately, rabies vaccines have become the most effective tool for preventing infection from this deadly viral zoonosis. Rabies vaccines can be used for both prevention and treatment, and current vaccines are more effective if administered promptly after exposure to rabies. Post-exposure prophylaxis (PEP), which includes cleaning the wound from RABV exposure, using rabies immune globulin (RIG) if necessary, and administering multiple doses of rabies vaccine, or pre-exposure prophylaxis (PrEP), which involves administering multiple doses of rabies vaccine before exposure (RABV), are the two main immunization schemes recommended by the World Health Organization (WHO) for preventing human rabies. The “Global Strategic Plan to Eliminate Human Rabies Deaths Caused by Dogs by 2030” was launched in 2018 by WHO, the World Organisation for Animal Health (WOAH), the Food and Agriculture Organization of the United Nations (FAO), and the Global Alliance for Rabies Control (GARC). It specifically emphasizes preventing canine rabies through annual mass vaccination, with at least 70% of dogs vaccinated.
More than a century has passed since Louis Pasteur developed the first vaccine for pre-exposure immunity and post-exposure prevention on June 6, 1885. Additionally, several rabies vaccines have been developed over the years and are now used to prevent or control rabies in humans and animals. Many countries around the world use cell culture-based inactivated rabies vaccines; however, these vaccines require multiple doses to elicit a strong humoral immune response and are more expensive to use for immunization in humans and animals in developing countries. Although inactivated neural tissue vaccines are relatively cheap, they have been eliminated in many countries due to their negative side effects, such as neurological paralysis complications in certain individuals. Modified live vaccines using SAD-Bern, Evelyn Rokitnicki Abelseth (ERA), and SAD-B19 strains can effectively induce protective immune responses with smaller amounts of virus, but there is still a possibility of causing rabies in animals due to residual virulence or pathogenic mutations during the virus’s replication in the host.
The further demand for safe, less reactive, and more immunogenic vaccines has led to the development of the next generation of vaccines. By introducing genetic manipulations in reverse genetics technology and recombinant DNA technology, our understanding of RABV biology has greatly improved. This has also significantly accelerated the creation of innovative vaccines, creating a platform for the next generation of vaccines. The initial genetically modified vaccines altered the rabies virus genome by deleting genes encoding phosphoprotein or matrix protein, resulting in non-pathogenic vaccine viruses that lack neurotoxicity even in immunocompromised mice. Transgenic vaccines, such as rERAG333E, ERAG3G, and SPBN GAS GAS strains, have a special status in inducing significant immune responses. For safety reasons, further modifications of recombinant vaccines have been made by manipulating the rabies virus genome to encode two copies of the glycoprotein or, in the case of nucleic acid vaccines, only express the rabies virus glycoprotein (RAVG), such as plasmids encoding the RABV G protein or mRNA-based rabies vaccines SFV-RVGP. However, these vaccines cannot maintain significant immune responses. The further development of less pathogenic viruses as RAVG carrier molecules has led to the emergence of viral vector vaccines, which provide a promising platform for oral vaccination of wildlife. Currently, viral vector-based vaccines RABORAL V-RG and ONRAB are favored in controlling wildlife rabies. Current research on the introduction of atypical rabies vaccination also represents a paradigm shift in post-exposure prevention in Asia.
This comprehensive review outlines the path traversed in the origins of rabies vaccines from mythological paradigms, and how this path has progressed in the practical history of rabies vaccines with the development of different types of conventional vaccines. Furthermore, this review emphasizes the emergence of current vaccines with advanced genetic technology and highlights the future focus of rabies vaccine research in eliminating rabies. Additionally, this review provides comprehensive information and highlights the various types of anti-rabies vaccines used in different countries, their advantages, limitations, success stories, and failures, which will enable us to design control strategies for the “Control of Dog-Mediated Human Rabies by 2030”.
02
The History of Rabies Vaccines
The history of rabies prevention began in the first century AD, with many myths and dogmas regarding rabies and its treatment. Until the nineteenth century, there was no consistent diagnosis or treatment for rabies in humans or animals. Techniques, including cauterization, were recommended for treating rabies wounds, and in some cases, they even involved amputation. However, all these beliefs never addressed the shocking death toll in humans and animals. In AD 25, people began to view rabies from a scientific perspective. Celsus in AD 25 promoted the early treatment of wounds after bites. In 1198, Maimonides depicted the long incubation period of bitten individuals. Later, Edward Jenner in 1769 identified the preference of the rabies virus for neural tissue. In 1804, Georg Gottfried Zink proved that the infectious saliva of rabid animals could be a source of infection. In 1852, French pharmacist Apollinaire Bouchardat became the first scientist to consider vaccination to prevent rabies. In 1881, French veterinarian Pierre-Victor Galtier achieved the first rabies immunization experiment in sheep by injecting rabies virus intravenously..
Although the history of vaccine development was pioneered by pre-Pasteur vaccine scientists, the actual history of rabies vaccine development began in 1885 with Louis Pasteur as an emergency management measure, even before the pathogen of the disease was identified..Initially, the etiology of rabies did not conform to Koch’s bacterial theory, as they could not cultivate any infectious agents related to the disease. Even in the late 19th century, rabies was believed to be caused by a parasite similar to a sporozoite..It was not until 1903 that they could prove any “filterable pathogenic agent” leading to this disease. The size of the rabies virus body was only determined in 1936, and the exploration of the pathogen using electron microscopy did not occur until 1962. Despite these limitations, in 1881, Pasteur and his team, along with Chamberland, Roux, and Thuillier, tracked the presence of the rabies virus in the central nervous system of animals suffering from rabies..
03
The First Generation Vaccine: Pasteur Vaccine (Neural Tissue Vaccine)
The era of the first-generation vaccine began with Louis Pasteur, who developed the first rabies vaccine extracted from the spinal cord of infected rabbits, physically inactivating the rabies virus by drying it. By adapting the virus’s characteristics in terms of virulence and incubation period through multiple passages and adaptation of street (wild-type) rabies virus to experimental animals, Pasteur was able to change the properties of the virus. By inoculating a stable amount of street virus preparation into the rabbit’s meninges and repeating the passage over 50 times, Pasteur observed a consistent latency fixed at 7 days from inoculation to the spread of rabies. Thus, he referred to this virus as the “fixed” virus. After several experiments on dogs as natural hosts, on July 6, 1885, Pasteur first injected his experimental rabies vaccine into a 9-year-old boy, Joseph Meister, who had sustained multiple severe rabid dog bites. About 2 days after the bites, the young boy received 13 injections of the dried rabies virus-infected rabbit spinal cord suspension, with increasing virulence, over 11 days. This strategic vaccination saved Meister from the brink of death from rabies..However, the main drawback of the Pasteur vaccine is that it contains increasingly virulent rabies virus. Moreover, there were concerns about the consistency of inactivation, as there were few reported cases of rabies after vaccination. Furthermore, the inability to produce enough vaccine to meet demand was the primary challenge in providing large-scale vaccine production. However, Pasteur’s method was subsequently used for over 50 years, after which significant improvements were introduced in the preparation of rabies vaccines.
04
Chemically Modified Fermi & Semple Vaccines
Through simple chemical modifications conducted by Fermi in 1908 and Semple in 1911, the Pasteur vaccine was further improved. The newer neural tissue vaccines (NTVs) were developed by Sir David Semple at the Central Research Institute (CRI) in Kasuli, India, from adult sheep (Semple vaccine). They used chemical agents, such as phenol, to inactivate the brains of infected sheep or goats..Although the addition of phenol inactivated the Pasteur vaccine, it still distorted the protein structure, compromising the immunogenicity of the rabies virus. Additionally, severe side effects, such as Guillain-Barré syndrome (GBS) and the risk of transmitting infectious spongiform encephalopathy, were reported. While this vaccine was widely used in many parts of the world, the WHO eventually suspended its use in almost all countries.
05
Non-Myelinated Tissue Vaccine
Although the Fermi and Semple vaccines were successful, the need for alternative, less reactive vaccines arose due to high myelin levels, sensitization in some vaccinated individuals, and the enhancement of sensitization leading to a few fatal encephalitis cases. In the 1940s, clinical studies on allergic meningoencephalitis and central nervous system demyelinating diseases associated with vaccination received significant attention. Later, the emergence of eggs containing embryos or the brains of newborn rodents, as a medium for producing rabies vaccines, made the journey of vaccine development safer. Clinical evidence showed the absence of substances leading to vaccine side effects in embryonic and newborn animal neural tissues. Researchers in the former Soviet Union developed a new rodent brain vaccine using mice..In 1964, Fuenzalida and his team developed a non-myelinated inactivated rabies vaccine from the brains of suckling mice (suckling mouse brain-SMB) through phenol inactivation and subsequent partial purification.
The lack of myelin in newborn animal tissues made the SMB vaccine less reactive compared to the Semple vaccine. However, studies indicated that this vaccine was not entirely free of myelin, and the presence of other undesirable components could lead to severe adverse reactions. Therefore, in line with WHO recommendations, global national regulatory authorities decided to cease the use of this vaccine after a decade of long-term use.
06
Embryonic Vaccine
In 1931, Ernest W. Goodpasture modified various human viruses using embryonated eggs, providing a new platform for further rabies vaccine development. After using live animals, embryonated eggs were employed to develop rabies vaccines. In 1940, the Flury strain of the rabies virus was used in 1-day-old chicks. This strain was subsequently established in chicken embryos. The Flury low egg passage (LEP) vaccine consisted of a modified live virus that underwent 40-50 generations of egg passage and was further lyophilized from a 33% whole embryo suspension. The measles vaccine was used for mass canine vaccination, but there were still some residual virulence, especially in kittens, cats, and cattle. Subsequently, the Flury high egg passage (HEP) vaccine was produced through a series of nearly 180 or more egg passages. Although this vaccine underwent human trials in the 1950s and 60s, it was eventually discontinued due to unsatisfactory vaccine potency. In the late 1950s, duck embryos became an alternative to chicken embryos in vaccine production, leading to the development of the duck embryo vaccine (DEV) for rabies, which contained rabies virus in a 10% whole embryo suspension, using β-propiolactone for inactivation. This vaccine was widely used for humans in the United States until the 1980s. Due to adverse reactions and poor immunogenicity, these vaccines were later discontinued. The successful adaptation of the rabies virus to embryos brought hope for alternatives to brain tissue vaccines. Although various strategic approaches have marginally improved the quality of these vaccines, questions regarding their safety, efficacy, and immunogenicity have not been fully resolved. Therefore, these vaccines were suspended in many regions globally, focusing on research into cell substrates for propagating rabies virus, leading to the era of cell culture vaccines.
07
Second Generation Vaccine: Cell Culture Vaccine
The establishment of cell culture systems for virus propagation led to a new paradigm in rabies vaccine development, resulting in the emergence of second-generation cell culture vaccines. Cell culture systems are a common method for producing viral vaccines because they offer many advantages over neural tissue vaccines and egg-based systems. They provide a defined safety and efficacy profile, reduce delivery time, and offer greater process flexibility. In 1930, rabies virus was cultured in primary explants of chicken embryonic brain cells and was further maintained for 5 consecutive passages..Later, research focused on the propagation of fixed RABV in mouse embryonic brain tissues. In 1942, Plotz and Reagan successfully isolated and cultured street rabies virus directly from the brains of rabid cases in primary explants of chicken embryonic cells for the first time..Subsequently, in 1958, the concept of culturing rabies virus in non-neuronal tissues was proposed, leading to the first tissue culture rabies vaccine using fixed rabies virus (from the brain of rabid mice) and street rabies virus (isolated from the salivary glands of rabid dogs) from primary hamster kidney cells. Kissling was able to continuously propagate the fixed virus through 15 cell culture passages and also successfully maintained the street virus through 4 passages. After that, in 1960, Fenje achieved the first adaptation of rabies virus strains using cell cultures that could be used for vaccine production, utilizing the street virus Alabama Dufferin (SAD) strain propagated in mouse brains and adapted through alternating passages in primary hamster kidney cultures and mouse brains..
Later, in 1963, Kissling and Reese prepared the first experimental cell culture rabies vaccine, the primary hamster kidney cell vaccine (PHKCV), from primary hamster kidney cells inoculated with previously adapted challenge virus standard (CVS) strain fixed rabies virus. In 1968, PHKCV developed using the fixed strain CL-60 (a derivative of the street virus Alabama Dufferin [SAD] rabies virus) was licensed in Canada..Later, in 1971, PHKCV was produced using the Vnukovo-32 strain from the former Soviet Union. Following Kissling, scientists began to adopt different cell culture systems to propagate various rabies virus strains for vaccine development. In 1964, Abelseth developed an attenuated rabies vaccine for livestock using the rabies virus SAD strain propagated in primary pig kidney cells..Later, in 1965, Kondo utilized the sensitivity of primary chicken embryonic cell cultures to propagate a Flury-HEP rabies virus strain adapted to egg cells, developing an inactivated rabies vaccine for humans..In 1969, Wiktor produced a purified concentrated rabies vaccine using the BHK-21 cell line derived from young hamster kidney cells. The rabies vaccine using bovine kidney cells from Pasteur virus (PV) and the dog kidney cell rabies vaccine using the Pitman-Moore strain (PM) were successfully developed in 1974 and 1978, respectively. Each of these received licenses for use in the Netherlands..
Through modern cell culture techniques utilizing diploid cell lines for vaccine production, the production of high-quality rabies vaccines has gradually become feasible. Subsequently, fixed RABV was grown in human diploid cell lines HDCS “WI 38,” developed by Hayflick and Moorhead in 1961. Later, in the mid-1970s, the WI-38 cell line was converted to the MRC-5 cell line for developing licensed human diploid cell vaccine (HDCV). HDCV was the first purified, concentrated, and lyophilized rabies vaccine without any adjuvants. Additionally, HDCV has fewer side effects. Therefore, the WHO recommends rabies HDCV as the gold standard reference vaccine, but the viral yield of human diploid cell vaccines is low and less economically feasible in many developing countries. This necessitated the development of alternative vaccines that are equally effective as human diploid cell vaccines, leading to the development of purified duck embryo cell vaccines and purified chicken embryo cell vaccines.
In 1971, the use of fully purified duck embryo-derived tissue vaccines improved through advanced purification schemes facilitated the production of purified duck embryo cell vaccines (PDEcv) using the CVS strain. In 1985, the Swiss Serum and Vaccine Research Institute approved the PDECV vaccine utilizing the Pitman-Moore strain. Additionally, PDECV is registered in certain European and Asian countries; however, it has not been authorized in the United States. These vaccines have proven superior to DEV as they are completely free of egg protein and myelin basic protein, which are significant sources of allergic meningoencephalitis. Later, in 1972, a purified chicken embryo cell vaccine (PCECV) was developed using the Flury HEP virus. Subsequently, another PCECV was developed and inactivated with the help of the Flury LEP virus, used as a vaccine for immunizing dogs for several years. The second human PCECV was developed by adapting the Flury LEP strain in chicken embryonic fibroblast (CEF) and was approved in Europe in 1984. In the United States, another PCECV was produced using the LEP-c25 virus and was licensed in 1997. Currently, PCECV is one of the most commonly used human rabies vaccines. Due to their considerable immunogenicity and tolerability, purified duck embryo cell vaccines (PDECVs) or purified chicken embryo cell vaccines (PCECVs) have become effective alternatives to human diploid cell vaccines for preventing human rabies in many parts of the world.
However, primary culture systems have inherent limitations on cell division. Diploid cell lines, such as WI-38, MRC-5, and FRhL-2, have a limited lifespan of approximately 50 consecutive passages, after which the cells become senescent. Although they have reproductive capabilities, the large-scale commercial culture adapted for vaccine production poses technical challenges. This has led to the use of continuous cell lines for vaccine production. In 1962, African green monkey kidney cells produced the Vero cell line. Compared to primary culture cells, Vero cells yield higher viral titers, simpler scaling for cell culture systems, and a long history of use in vaccines without causing safety issues. In the early 1980s, rabies virus was propagated in Vero cells for vaccine production. Vero cells support the replication of various rabies viruses, including HEP, CVS, Mokola virus, Duvenhage bat virus, and Lagos bat virus. The advantages of Vero cells have significantly reduced the cost of producing rabies vaccines, enabling access to rabies vaccines for most developing countries. The further introduction of purified Vero cell-derived rabies vaccines (PVRV) into clinical practice remains crucial for rabies prevention. In 1985, purified Vero cell rabies vaccine (PVRV) was licensed in Europe. Furthermore, the WHO proposed replacing neural tissue vaccines with more effective and safer vaccines developed through cell culture..Currently, PVRV is widely used globally. As an advancement in the vaccine production process, a next-generation enhanced serum-free PVRV vaccine was produced from inactivated rabbit plague virus PM strain..This next-generation vaccine’s immunogenicity and safety make it a new option for rabies prevention.
08
Recent Advances in Rabies Vaccines
Although many anti-rabies vaccines have been invented over the years to protect humans and animals from rabies, the safety and immunogenicity of candidate vaccines remain paramount. However, the fundamental concepts of vaccine development, such as attenuation and inactivation, have consistently been the pillars of ongoing rabies vaccine innovation. Ongoing efforts in rabies vaccine research primarily focus on enhancing the immunogenicity and safety of candidate vaccines, leading to the emergence of improved or inactivated vaccines.
8.1. Modified Live Vaccines (MLV)
Modified live vaccines are typically produced by altering the pathogenicity of naturally occurring viruses, allowing them to elicit a robust immune response without causing clinical disease. Most scientists modify the virus through continuous passages in different cells to ensure the future safety of the vaccine, leading to the development of attenuated live vaccines for various diseases, including rabies. The original SAD (street virus Alabama Dufferin) strain of the rabies virus is the source of all currently used attenuated vaccines, which have undergone multiple passages in cell cultures.
In some Asian countries, a chicken embryo-derived MLV vaccine of the Flury strain has been developed and used in animals. Meanwhile, MLV vaccines have been produced using the street virus-Alabama-Dufferin (SAD) strain for further use. To create a vaccine of superior quality to the Flury-LEP vaccine, a modified live vaccine strain, Evelyn-Rokitnicki-Abelseth (ERA), was introduced in Canada in 1974, as the Flury low egg passage vaccine had negative consequences due to tissue fragments in the vaccine. Ultimately, the Flury low egg passage vaccine was replaced by the ERA strain by the Korean Veterinary Bureau in the late 1970s. Livestock (including dogs, lambs, goats, and cats) vaccinated with the ERA attenuated live vaccine via intramuscular routes exhibited no clinical symptoms, and no vaccine strains were recovered from their salivary glands or brains, yet it still elicited a strong immune response and elevated levels of virus-neutralizing antibodies (VNA). Unfortunately, nearly 50% of dogs receiving intracranial injections of the vaccine exhibited significant clinical symptoms, such as anorexia, fever, severe tremors, mild paralysis, and paralysis. Due to their residual virulence, this remains the main drawback of modified live vaccines. Additionally, MLVs are more sensitive to temperature fluctuations, and accidental self-vaccination with MLV rabies vaccines poses a risk to vaccinators..
8.2. Inactivated Rabies Vaccines
The history of inactivated vaccines began about a century ago when inactivated neural tissue vaccines developed for rabies were used in certain African and Asian countries. Most traditional rabies vaccines use fully inactivated viruses with the same antigenic characteristics as wild-type viruses. It has been shown that immunization with fully inactivated viruses activates helper and cytotoxic T cells and defends against fatal central nervous system rabies virus attacks, leading to the production of virus-neutralizing antibodies.
Globally, various rabies virus strains have been used to produce inactivated rabies vaccines, including CVS 11, Pittman-Moore-NIL2, RC-HL (produced from the Xiyuan strain), and Pasteur virus strains. Currently, rabies vaccines approved for humans are based on inactivated purified rabies viruses propagated in cell cultures or embryonated duck or chicken egg systems. Rabies vaccine strains are typically inactivated using β-propiolactone (BPL), ultraviolet light, acetylamide, or ethyleneimine (BEI). BPL is the most commonly used inactivating agent; however, it is expensive and unstable at 37°C. Due to phenol and formaldehyde potentially distorting the structure of antigenic sites, their use for viral inactivation is no longer recommended. In contrast, BEI is less hazardous to handle and has advantages of good stability, low cost, and ease of preparation. However, the fundamental drawbacks of inactivated vaccines are their lower immunogenicity, expense, and the need for multiple dosing regimens, especially in pre-exposure and post-exposure immunization..
8.3. Adjuvanted Rabies Vaccines
Adjuvants are substances that enhance or modulate the immune response to vaccines by increasing the inflammatory response necessary for original B and T cell stimulation driven by the antigen. Adjuvants have been a focus of attention due to the use of inactivated vaccines, subunit vaccines, and synthetic vaccines, which are fundamentally weakly immunogenic, and adjuvants are used as supplements to enhance immunogenicity and produce more effective vaccines due to their immunoamplifying capabilities. Common adjuvants include aluminum hydroxide, aluminum phosphate, and saponins..However, most adjuvants currently in use are aluminum salts. Although alum was the first adjuvant approved for humans, it is claimed to delay the early production of antibodies and is not equally effective in inducing cellular immunity. The main drawbacks of adjuvants are the adverse side effects, toxicity, and limited adjuvanticity of certain formulations against a few antigens, necessitating the development of synthetic derivatives, such as peptidoglycan, liposomes, QS21, monophosphoryl lipid A, MF-59, and immunostimulating complexes (ISCOMS) as alternative adjuvants. In recent years, many studies have focused on developing alternative adjuvants to improve the immunogenicity and efficacy of inactivated rabies virus vaccines, and various other compounds, including indigo root polysaccharides, CpG oligodeoxynucleotides, monophosphoryl lipid A (MPLA), β-glucans, hyaluronic acid derived from Staphylococcus aureus, and purified protein derivatives (PPD) from BCG, have been developed. These candidate adjuvants are undoubtedly promising platforms for developing new adjuvanted rabies vaccines with enhanced immunoenhancing potential and reduced risks of adverse health impacts.
09
Next-Generation Vaccines
The risks posed by the residual virulence of modified live vaccines and the low immunogenicity of inactivated vaccines, along with the large amounts of antigens and repeated doses required to elicit protective immune responses, ensure the demand for safer and more economical rabies vaccines. Currently, the emergence of technologies such as recombinant DNA technology and reverse genetics has provided in-depth insights into the rabies virus genome and facilitated genomic manipulation, which in turn brings greater hope for more effective and safer vaccines, reducing costs, and improving stability and immunogenicity. This has led to the era of next-generation vaccines targeting recombinant rabies virus strains or individual recombinant rabies glycoproteins (G proteins), which help overcome the drawbacks of attenuated live vaccines.
9.1. Transgenic Vaccines
Genetically modified viruses are produced by genetically modifying the viral genome through biotechnological methods, involving targeted insertions, deletions, artificial synthesis, or modifications of nucleotide sequences while retaining infectivity. Most rabies vaccines involve some form of attenuation, weakening, or inactivation of the virus to render its toxic characteristics ineffective. Genetic focus on the viral genome has confirmed that the glycoprotein (G) is most closely related to RABV pathogenicity, and certain amino acid sites associated with viral pathogenicity have been identified. Therefore, generating site-specific mutations or insertions in these amino acids concerning the parent rabies virus strain will eliminate residual pathogenicity, prevent potential virulence reversion, reduce the likelihood of reversion to potential original amino acids, and enhance the safety of the resulting mutants, which in turn will be a promising option for producing highly attenuated rabies vaccines.
In particular, these vaccines are crucial for mass immunization of stray dogs in the habitats of wildlife in both developing and developed countries, and as a result of various research efforts, many mutant vaccine strains have been developed. A genetically modified ERA vaccine strain with an arginine to glutamic acid mutation at residue 333 of the RVG (G333E) has been developed. This live rERAG333E can induce strong and persistent RVNA in dogs through intramuscular injection and oral immunization in mice, with an intensity higher than that of the ERA strain wild type (rERA)..Furthermore, using reverse genetics, the complete genome of rRABV was mutated at amino acid position 333 of the RVG gene, and auxiliary plasmids were rescued in BHK/T7–9 cells, leading to the construction of the ERAG3G strain, which showed complete protection against pathogenic RABV after further oral immunization in mice. Another new strain, ERAGS, is a rRAVB with site-specific mutations at positions 194 and 333 of RVG, found to be non-pathogenic and extremely safe in immunized mice against highly pathogenic RABV. A new and highly attenuated bivalent rabies virus construct, SPBN GASGAS, was produced from the rabies virus strain SAD L16 by mutations at amino acid positions 194 and 333 and an additional identical changing glycoprotein gene. By reducing the potential risk of reversion to virulence and enhancing apoptosis, the resulting mutant exhibits improved safety. Recently, SPBN gas has occupied a special position as an effective rabies vaccine candidate for oral vaccination to mitigate rabies in free-roaming dogs in certain developing countries (such as Thailand, Haiti, Namibia, and Morocco).
9.2. Recombinant Rabies Vaccines
For safety and to enhance immunogenicity, further recombinant vaccine variants have been produced by editing the rabies virus genome to encode two or more copies of the glycoprotein or using strategies that solely clone and express the rabies virus glycoprotein (RAVG). The recombinant rabies vaccine constructs encoding two copies of the glycoprotein gene provide better protection in mice and protection against the virulent challenge virus standard-11 (CVS-11) strain in dogs..These studies have also revealed that RABV containing two copies of the G gene increases the expression of the G gene, significantly enhancing the vaccine’s potency, boosting immunogenicity, producing higher levels of virus-neutralizing antibodies, and reducing pathogenicity. The increased expression of the G gene is associated with enhanced apoptosis, which helps induce a strong upregulation of genes related to host immune responses, observed in neurons infected with attenuated RABV strains..Therefore, these recombinant RABV strains may be promising candidates for future inactivated vaccines.
9.3. Nucleic Acid-Based Rabies Vaccines
The genetic material from pathogenic viruses is used in nucleic acid vaccines to produce protective immune responses against infectious materials. Nucleic acid-based vaccines have the potential to be cost-effective, safe, and efficient. Moreover, the immune response induced by nucleic acid vaccines focuses solely on selected pathogen antigens. Nucleic acid vaccines can be DNA (e.g., plasmids) or RNA (e.g., messenger RNA (mRNA)) and have shown significant potential for treating various diseases..This is based on cloning DNA into delivery plasmids or expressing antigens in host cells through direct mRNA (mRNA) inoculation. Additionally, host mechanisms will facilitate the synthesis of endogenous proteins, simulating natural infections that trigger both cellular and humoral responses against the expressed antigens.
9.3.1. Rabies DNA Vaccines
Typically, sufficient levels of virus-neutralizing antibodies against the rabies virus glycoprotein are directly related to rabies protection. Recombinant DNA technology can easily clone the glycoprotein gene into appropriate expression vectors, facilitating effective expression of the glycoprotein in vivo and leading to the production of rabies DNA vaccines. Since 1994, DNA vaccines have been regarded as a more economical and effective method for rabies prevention, with their feasibility confirmed in various animal models, including companion animals..The glycoprotein sequences of Pasteur virus (PV), challenge virus standard (CVS), Evelyn Rokitnicki-Abelseth (ERA), or street virus isolates have been used to evaluate DNA rabies vaccines. The RV glycoprotein has been expressed in various expression vectors, including pSG5, pCIneo, pVR105, DNAVACC, and has successfully induced specific immune responses in experimental animals. Although this technology has proven effective, its widespread use, especially in post-exposure prevention for rabies, has been hindered by slower and weaker immune responses. However, the co-administration of glycoprotein-encoding plasmids with chemical adjuvants or the co-delivery of cytokine genes and innovative delivery technologies, including electroporation after DNA injection and co-administration with traditional inactivated rabies vaccines, have proven effective in enhancing immune responses and increasing their potential for preventing and managing rabies. Furthermore, current developments in vector design and delivery systems also bring hope for improving the effectiveness of rabies DNA vaccines. However, the need to address the risks of integrating plasmid DNA into host chromosomes, the formation of tolerance to plasmid DNA vectors, and antigen and autoimmune-related risks must be critically resolved before they can be developed into widely used vaccines.
9.3.2. Rabies RNA Vaccines
RNA vaccines are the simplest type of nucleic acid vaccine and a highly promising alternative platform for vaccine development. RNA vaccines have many advantages over DNA vaccines. RNA vaccines are translated directly in the cytoplasm, eliminating the need for transport to the cell nucleus, leading to rapid antigen expression without the risk of possible integration into the host genome. RNA is known to be a potent adjuvant, stimulating immune responses through pattern recognition receptors, such as Toll-like receptors or retinoic acid-inducible gene I-like receptors. The self-replicating ability of messenger RNA (mRNA) has led to the development of two main types of RNA vaccines: traditional non-amplifying mRNA vaccines and self-amplifying mRNA vaccines from RNA viral vectors (also known as replicons), such as Semliki Forest virus (SFV), which retains replicative activity. Studies have shown that traditional non-amplifying mRNA-based rabies vaccines encoding the RABV G gene stimulate effective VNAs and antigen-specific CD4+ and CD8+ T lymphocytes in immunized mice, protecting them from lethal central nervous system infection..Similarly, mRNA-based rabies vaccines have shown immunogenicity in pigs with effective VNA levels. A self-amplifying mRNA rabies vaccine containing recombinant SFV encoding the RABV G protein (SFV-RVGP) can induce high levels of functional trimeric RABV G protein expression, eliciting antibody levels comparable to commercial rabies vaccines, and is more effective than protein vaccines in generating cellular immune responses..However, a major challenge is the instability of RNA vaccines due to RNase-mediated degradation or electrostatic repulsion due to interactions between negatively charged mRNA molecules and negatively charged cell membranes, leading to transient expression of antigens after RNA delivery, which requires heightened attention.
9.4. Rabies Protein Subunit and Peptide Vaccines
Protein vaccines stimulate the immune system by using peptides or proteins as antigens. Protein antigens can be naturally derived from pathogens causing infectious diseases and can be in their complete form or derived from lytic products. Protein subunit vaccines can also be produced as heterologous proteins in recombinant systems, including recombinant bacteria, yeast, insect cells, or mammalian cells, as alternatives to natural sources. The RABV G protein is a major determinant of the virus’s pathogenicity and is the primary protective antigen responsible for inducing protective immunity against rabies. Therefore, the efficacy of G protein (purified from infected cells) has been established in various systems and has been used as a form of protein vaccine for preventing rabies. However, these methods have proven ineffective. Yeast-derived proteins failed to elicit protective immune responses in mice, likely due to poor folding of the end products. The G protein produced from baculovirus in insect cells has been shown to be immunogenic, but the extensive purification required may render this method uneconomical. Currently, the production of vaccines using transgenic plants is gaining increasing importance. Researchers have also utilized plants, such as tomatoes, corn, cantaloupe, and tobacco, to express RABV G protein to create edible vaccines..This vaccine can produce detectable VNA responses in mice after ingestion and provide protection against challenges. Although these types of vaccines have the potential for low-cost production, several obvious drawbacks exist, including immunogenicity, stability of the vaccine in fruits, degradation in the stomach, and intestinal immune responses. Therefore, while edible vaccines represent an ideal option, further research is needed before they can be considered a viable alternative to rabies vaccines.
In addition to protein vaccines, many antigenic epitopes of the RABV G protein have been identified, and synthetic peptides mimicking these G protein epitopes have been produced and used as vaccines. Animals vaccinated with synthetic peptides encapsulating the rabies virus G5 antigenic region produced antibodies with strong affinity but low neutralizing potential. The rabies virus glycoprotein peptide mimetic epitopes (RABVG site III mimetic epitope (C-KRDSTW-C)) demonstrated stronger immunogenicity in mice and provide a new concept for future rabies vaccines..However, vaccines made from proteins or peptides exhibit moderate immunogenicity; the 65kDa viral glycoprotein is synthesized, forming trimers, and is moderately N-glycosylated at one of three possible sites. The rabies virus glycoprotein must be correctly folded into its native trimeric form to produce neutralizing antibodies, but this remains a challenge for protein vaccines.
9.5. Gastrointestinal Virus Vector Rabies Vaccines
Viral vector vaccines have been developed by cloning the desired antigen into heterologous viruses, which serve as carrier molecules to transfer the foreign gene into host cells, leading to its subsequent expression. The first viral vector expressing foreign genes was produced in 1972 from simian virus 40; since then, various viruses, including adenoviruses, poxviruses, herpesviruses, vesicular stomatitis virus, and lentiviruses, have been genetically modified to be non-pathogenic and repurposed as vaccine vectors, potentially stimulating the immune system against the proteins produced by the encoded transgenes. The antigen-specific cellular immune responses and strong and durable antibody levels induced by viral vectors, as well as their inherent adjuvanticity, make them more effective candidates in rabies vaccine development.
Several researchers have targeted different viral vectors to express rabies virus glycoprotein. RVG vaccination using recombinant adeno-associated viruses (AAVs) enhanced the protective levels of rabies virus neutralizing antibodies (RVNAs) after a single intramuscular vaccination..Several adenoviruses (Ads) have been widely used as vaccine vectors in vaccine development because they elicit strong humoral and cellular immune responses. In particular, trials of rabies vaccines using simian Ad vectors expressing the rabies virus glycoprotein have been conducted. A chimpanzee adenovirus vector rabies virus glycoprotein cloned AdC68-rab was shown to significantly elevate RVNA titers in mice four weeks after muscle injection..After intramuscular vaccination, this vector induced high levels of RVNA titers and more effective cellular immunity in mice, as well as very rapid, sustained, robust, and durable RVNA levels in rabbits, with protective levels lasting up to 48 weeks in macaques. These effects were comparable to those induced by one, two, or even three doses of the Rabipur vaccine. Compared to other Ad serotypes, ChAdOx2 RabG with chimpanzee Ad serotype 68 (AdC68) and human Ad serotype 5 (AdHu5) exhibited higher immunogenicity and induced high levels of RVNAs in vivo with a single IM dose. The protective immune responses generated by the chicken adenovirus rabies virus glycoprotein (CAV2-RVG) through intramuscular, intranasal, and oral immunization in mice and dogs are quite remarkable. Additionally, other viral vectors, including parapoxvirus Orf virus (ORFV), vesicular stomatitis virus (VSV), raccoon poxvirus (RCN), single-cycle yellow fever virus, Newcastle disease virus (NDV), and bovine herpesvirus type I (BHV-1), express rabies virus glycoprotein and effectively induce RVNAs, maintaining their status as effective vaccine candidates. Nevertheless, a common obstacle with viral vectors is the pre-existing immunity in hosts against certain vectors, which may minimize vaccine efficacy. However, this can be avoided by using alternative vectors with low seroprevalence in the host. Furthermore, the development of viral vector rabies vaccines has progressed, becoming oral rabies vaccine candidates and fundamental tools to reduce rabies virus mortality in global wildlife species.
10
Oral Rabies Vaccines
Animal vaccines are an effective method for protecting public health by reducing the pathogens of animal infectious diseases. The long history of rabies vaccine development has provided many effective vaccines for parenteral vaccination strategies. However, vaccinating wildlife or free-ranging livestock (such as pet dogs) through parenteral vaccination remains a challenge. Therefore, the demand for alternative vaccination strategies to eradicate rabies has led to the development of oral rabies vaccines (ORV). Oral vaccines use bait for oral administration, which is designed with sealed pouches containing vaccine suspension encapsulated in palatable bait materials, such as animal intestines, chicken heads, eggs, and dog food-based materials, depending on the target species. When consumed, chewing actions perforate the pouches, releasing the vaccine suspension into the mouth. Here, the vaccine is primarily absorbed by the palatine tonsils, and after a small replication at the entry site, it triggers a protective immune response. Currently, there are two commercially licensed ORVs available for wildlife rabies immunization. They are modified live vaccines (MLV) and vector-based vaccines (VBV). In modified live vaccines, the rabies virus is modified to weaken its virulence but still has the ability to trigger the immune system to produce antibodies. In vector-based vaccines, the gene encoding the antigenic glycoprotein is inserted into a viral vector, which can induce an immune response by expressing the inserted rabies virus glycoprotein during immunization. Since 1978, oral rabies vaccines (ORV) have been successfully used to control wildlife rabies in Europe and the United States.
10.1. Modified Live Virus Oral Rabies Vaccines
The development of modified live rabies virus oral vaccines began in 1935, derived from the street virus Alabama Dufferin (SAD) rabies virus strain. This parental SAD strain was isolated from the salivary glands of rabid dogs in the United States. Subsequently, the SAD strain underwent continuous passages in non-natural hosts, such as mice, chicken embryos, and non-neural cell lines, including hamster kidney and pig kidney cells, to enhance its heat stability, resulting in highly attenuated strains SAD-Bern, Evelyn Rokitnicki Abelseth (ERA), and SAD-B19. These strains constitute the first generation of ORV, which is the cornerstone of controlling wildlife rabies in Europe and has been widely used globally..Moreover, to improve the safety of the first generation of ORV, monoclonal antibodies were used to induce selected mutations in the rabies virus SAD strain, producing second-generation ORVs, SAD VA1, SAG1, or SAG2 strains. In 1990, a double mutant strain SAG2 was isolated from the rabies virus SAD-Bern strain, affecting two consecutive mutations at the glycoprotein’s 333 amino acid residue. Due to its safety and genetic stability, the SAG2 strain has proven to be an improved version of SAG1. Although these vaccines successfully reduced rabies levels in wildlife, there remains a need to effectively control rabies in large populations of free-ranging dogs in most rabies-endemic countries. This necessitates a safer candidate vaccine to supplement parenteral vaccination in large-scale canine vaccination activities, which remains crucial.
In certain developing countries, successful field trials of immunizing free-roaming dogs using SPBN gas have been significant. A vaccination campaign using bait containing SPBN gas was conducted on 2444 dogs in Thailand, with approximately 65.6% of stray dogs effectively receiving oral immunization. In Haiti, large-scale vaccination of dogs in field conditions was assessed, showing that 78% of dogs produced immune responses after consuming the bait. Monitoring seroconversion immune responses for up to 56 days post-vaccination in locally free-ranging dogs in Namibia demonstrated the immunogenicity of the highly attenuated vaccine strain SPBN GASGAS, successfully protecting approximately 79% of dogs with oral vaccination. These investigations confirm the immunogenicity of the SPBN GASGAS vaccine strain and the capacity of ORV to reduce dog-mediated rabies in developing countries. Furthermore, the feasibility of using oral bait for rabies vaccination in stray dogs in Goa, India, has been experimentally demonstrated, supporting the notion that ORV will be a promising method to significantly improve dog vaccination coverage in the country.
10.2. Vector-Based Oral Rabies Vaccines
The development of vector-based vaccines helps overcome the risks associated with MLVs. These vector-based rabies vaccines hold promise for veterinary and human use. Several research efforts have produced effective vector-based ORVs. Recombinant vaccinia viruses expressing the rabies virus glycoprotein have been tested as oral rabies vaccines in bait and have demonstrated the induction of protective immunity in several wildlife species. In vaccine development, various adenoviruses have been widely used as vaccine vectors due to their ability to elicit strong immune responses. The development of oral vaccine baits based on the replicating human adenovirus type 5 (AdHu5) expressing rabies glycoprotein (ONRAB) has been encouraging in controlling rabies in North American wildlife. The efficacy and safety of first-generation replication-defective adenoviral vectors have improved compared to second-generation replicating adenoviral vectors. Some reports indicate that pre-existing immunity against AdHu5, detected in 45-90% of the population, inhibits the immune response induced by AdHu5 vectors. To overcome these issues, rare serotypes of human adenoviruses or adenoviruses from non-human species, such as chimpanzees and canids, have been utilized for vector construction and vaccine development. The vaccine based on the recombinant chimpanzee adenovirus serotype 68 expressing the rabies virus glycoprotein (AdC68) has induced effective VNAs through intranasal or oral administration in newborn mice..Orally, this vaccine shows promising effects in young, pre-exposed individuals. Furthermore, rabies vaccines based on chimpanzee adenovirus vectors have shown protection in dogs against lethal rabies virus attacks. Studies have documented protective immune responses generated through oral immunization in mice and dogs using canine adenovirus type 2 rabies virus glycoprotein (CAV2-RVG).
Currently, there are two commercially licensed VBVs available for controlling wildlife rabies. RABORAL V-RG, which uses a recombinant vaccinia virus vector, and ONRAB, which uses a recombinant human adenovirus vector. One of the main issues with using VBVs is the potential infections caused by the vector viruses themselves. Research emphasizes that exposure to recombinant vaccinia virus vector vaccines in the United States may lead to severe skin inflammation in humans and complications in pregnant and immunocompromised individuals; however, no public reports have been made regarding adenoviral vectors. Additionally, the potential interference of pre-existing immunity against the vectors is a major concern with vector vaccines, as it prevents the generation of adequate immune responses against rabies. Nevertheless, the current viral vectors have been modified to evade pre-existing immunity and make them more like a potential delivery system. The success stories of oral vaccines in effectively controlling wildlife rabies and their potential to induce immunity in experimental studies highlight the effectiveness of oral vaccines as a supplementary strategy for controlling rabies in developing countries.
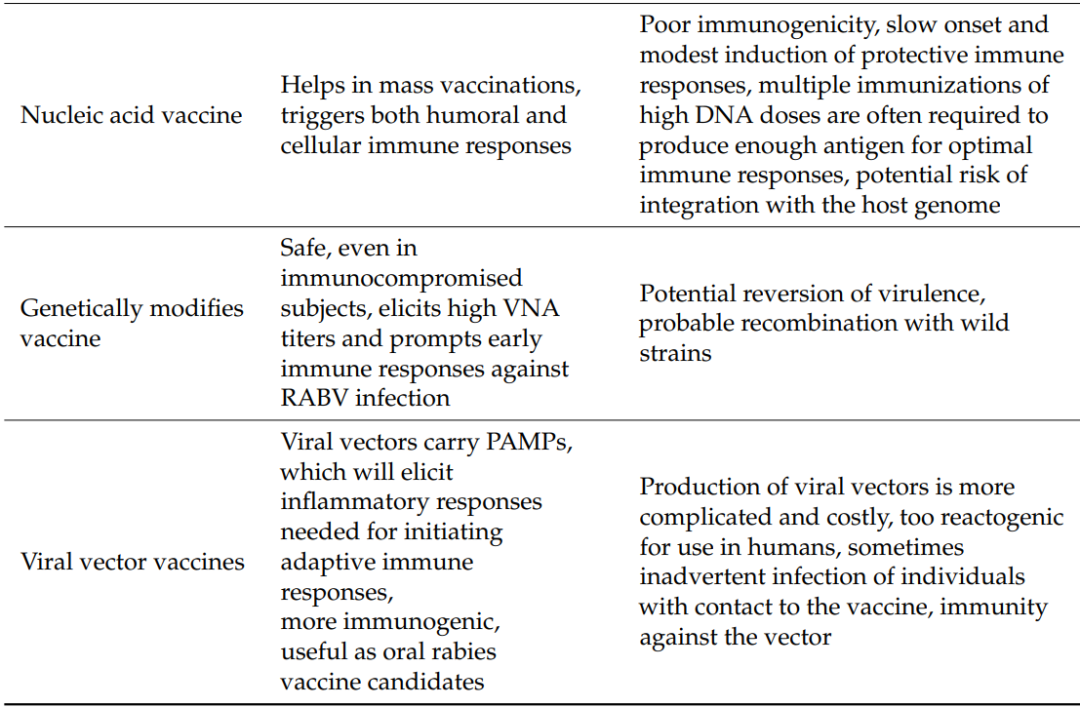
Details of the current research on different vaccine developments and the advantages and disadvantages of various vaccines are summarized in Table 1 and Table 2.
Table 1. Current Status of Rabies Vaccine Research
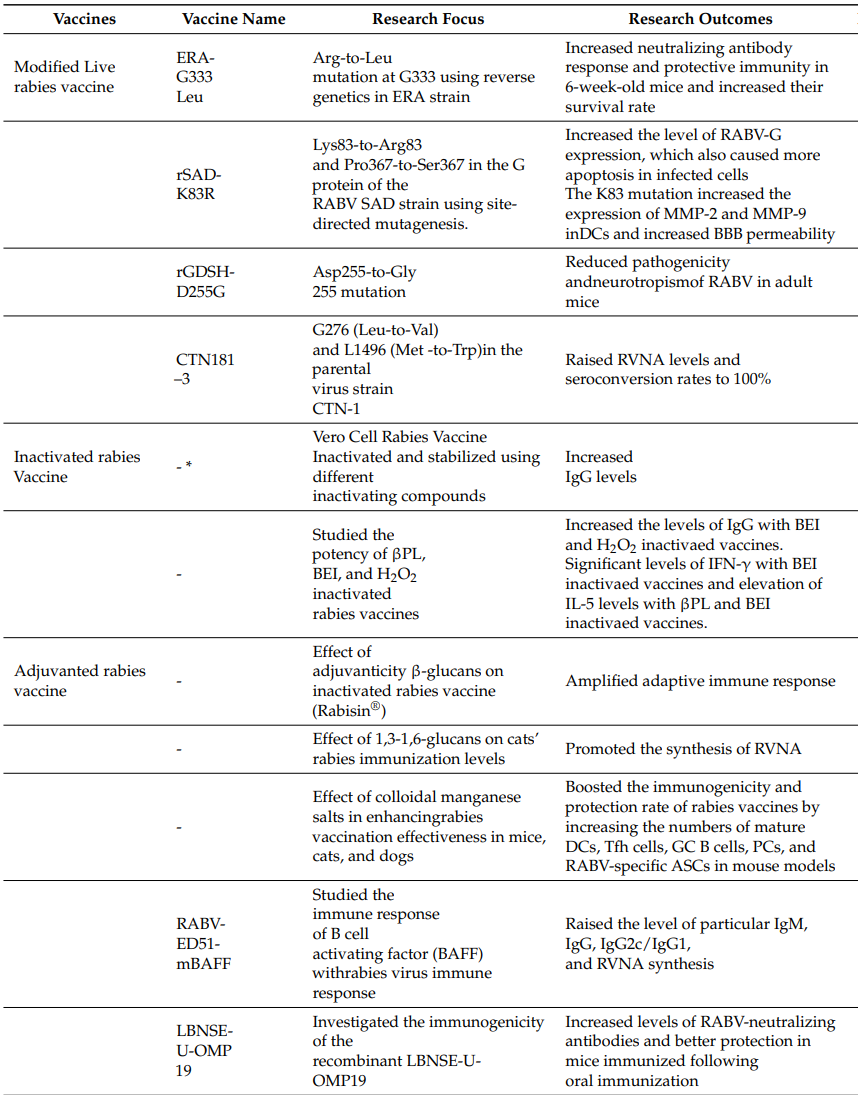
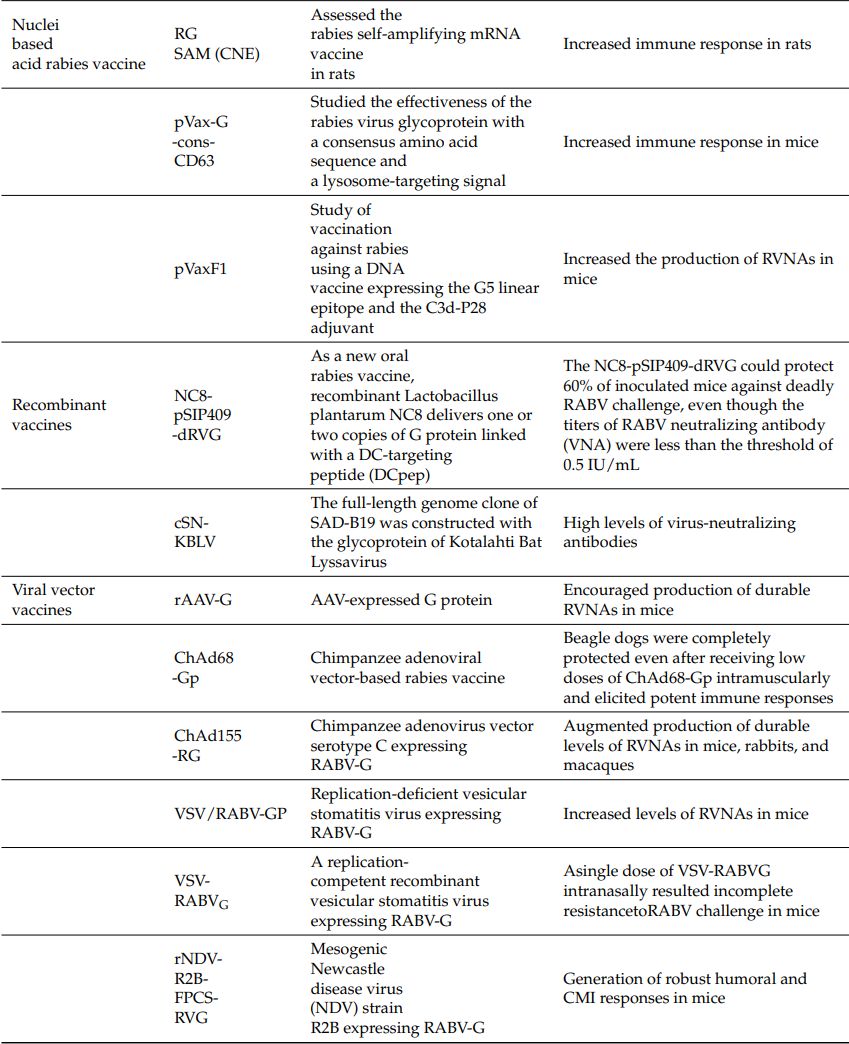
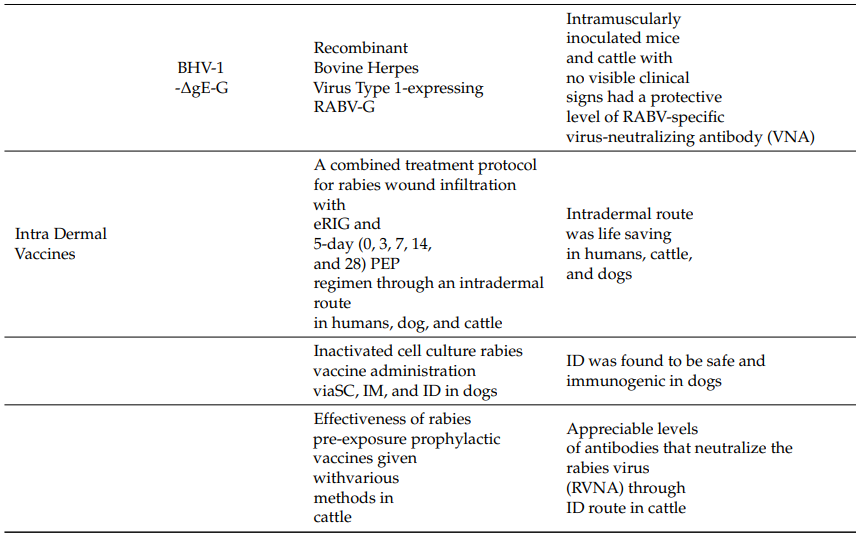
* Vaccine names unspecified
Table 2. Advantages and Disadvantages of Current Rabies Vaccines
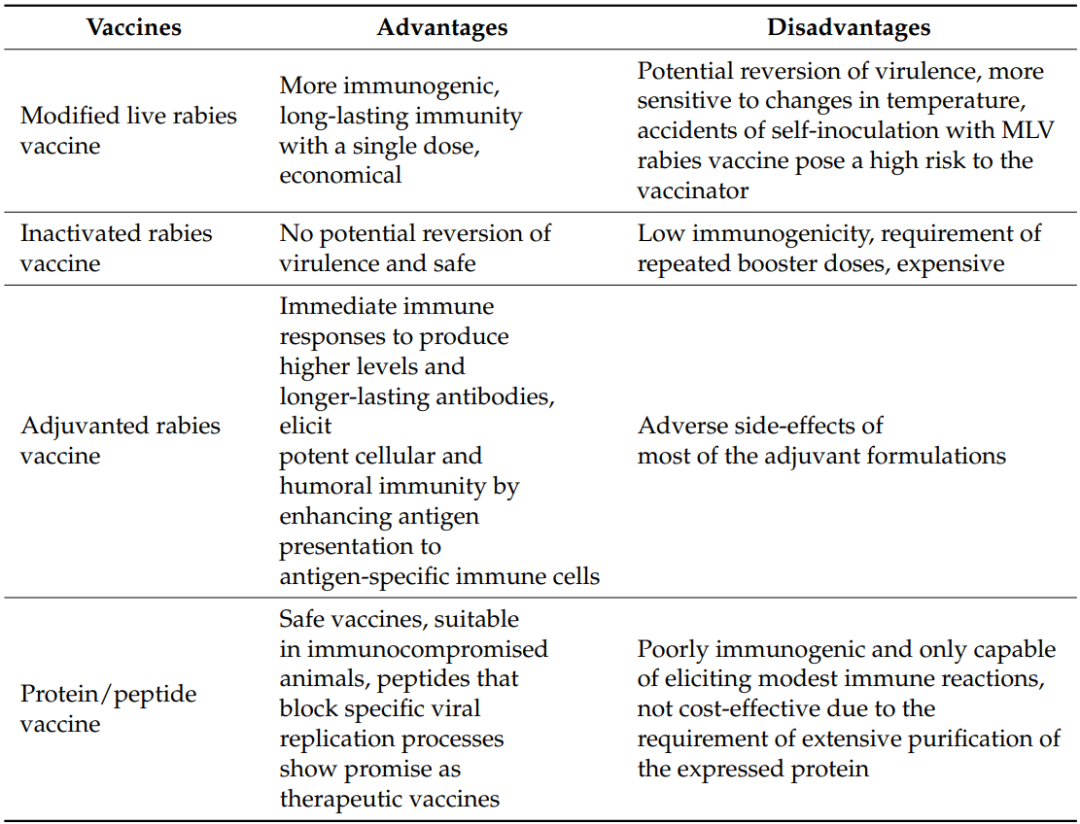
11
Intradermal Rabies Vaccine Administration
The ability of a vaccine to stimulate an immune response also depends on the route of vaccine administration. Intradermal administration delivers the antigen to the space between the epidermis and dermis. This space is anatomically favorable for immune stimulation, as the dermis and epidermal layers are rich sources of antigen-presenting cells, such as Langerhans cells, dermal dendritic cells, and dermal macrophages, known to participate in vaccine-induced immune responses, and the skin is regarded as an ideal target for vaccination. The extensive network of capillaries and lymphatics in the dermis also facilitates the movement of leukocytes and dendritic cells from the skin to secondary lymphoid organs. Most rabies vaccines are administered intramuscularly (IM) in increased amounts of antigens. Therefore, due to the cost of these antigens and their occasional scarcity, its use is limited in many underdeveloped countries. Administering reduced doses (typically 10% or 20% of the standard antigen dose) through the intradermal (ID) route can elicit immune responses similar to those caused by the standard doses administered via the IM route.
In the United States, the intradermal method for pre-exposure immunization with rabies vaccine was first approved in 1986, followed by the development of a cost-effective multi-site intradermal (ID) immunization method. The WHO Expert Committee recommended the use of modern rabies vaccines for post-exposure prevention via intradermal injection in 1991. According to a WHO review, cell culture rabies vaccines administered through the ID route (with an efficacy of > 2.5 IU per intramuscular dose) have the same or higher efficacy as those administered via the IM route for post-exposure prophylaxis (PEP) or PrEP. Similarly, the efficacy of rabies pre-exposure prophylactic vaccines administered through various methods in cattle revealed considerable levels of antibodies neutralizing rabies virus (RVNA) through ID administration. The combined treatment regimen using eRIG and intradermal injection over 5 days (0, 3, 7, 14, and 28 days) for rabies wound infiltration can save the lives of humans, cattle, and dogs, while the intradermal route has proven to be more economical due to its dose-saving effects. Similarly, the administration of inactivated cell culture rabies vaccines via subcutaneous, intramuscular, and intravenous routes has been found to be safe and immunogenic. These clinical trials provide confidence for promoting ID-administered rabies vaccines in regions where PEP cannot be accessed due to cost, and ID as a cost-saving and dose-sparing alternative immunization strategy should be encouraged.
12
Rabies Virus Immunity: Natural Infection vs. Rabies Vaccination
12.1. Immunity to Natural Rabies Virus Infection, Including Humoral and Cellular Responses
RABV typically infects the nervous system (NS). However, in the early stages of infection, viral particles are “injected” into the muscle and skin, potentially triggering immune responses in the periphery before the virus enters the nervous system. Once RABV enters the NS, it is less likely to elicit primary adaptive immune responses since the NS is an immune-privileged site, lacking specialized antigen-presenting cells and lymphoid tissues. However, after entering the NS, RABV induces early innate immune responses characterized by antiviral, chemotactic, and inflammatory responses, all involving infected neurons. Due to insufficient immune efficacy in the NS, RABV employs immune evasion strategies to escape host immune responses, successfully adapting to the host system, and cell-mediated immune (CMI) responses play a crucial role in blocking viral invasion and restraining RABV from entering the peripheral nervous system (PNS) during the early stages of infection. However, the blood-brain barrier poses a significant barrier to CMI responses. Additionally, the expression of HLA-G molecules in neuronal cells through the production of interferon-β leads to the conversion of naive human T cells into regulatory T cells, and the production of inhibitory cytokines such as interleukin-10 leads to cellular immune suppression..Moreover, the humoral immune system is a key defense mechanism against the rabies virus. Neutralizing antibodies can block the infectivity of the virus and prevent it from adhering to the neuronal cells of immunized individuals. Research indicates that unvaccinated animals cannot produce optimal levels of neutralizing antibodies when exposed to RABV, allowing the virus to reach the nervous system and manifest as disease..
12.2. Immunity from Rabies Vaccination, Including Humoral and Cellular Responses
Rabies vaccines’ most important function is to induce sustained antibody responses by activating CD4+ T lymphocytes. While cytotoxic T cells are generally considered more effective at clearing viral infections from tissues than antibodies, rabies is an exception. Additionally, the activation of CD8+ T cells leads to clinically associated pathogenic responses related to paralysis. Given the high risk of strong detrimental CD8 responses generated in the nervous system, this knowledge typically prevents the use of live vaccines, such as DNA vaccines or recombinant viruses, as post-exposure immunization. However, due to the effectiveness of live immunization, these next-generation vaccines remain applicable to pre-exposure vaccination schemes.Maintaining the integrity of the nervous system is best achieved with inactivated post-exposure vaccines, which primarily promote B cell activation with the help of CD4+ T cells. Because post-exposure immunization establishes immune responses in peripheral secondary organs, they are likely to provide protection. Activated lymphocytes, CD4–producing plasma cells, secrete antibodies into the nervous tissue to neutralize the virus..13
Methods of Treating Rabies with Vaccines and Antibodies
13.1. Rabies Vaccine Therapy
Thousands of people die from rabies each year, most of which occur in Asia and Africa. However, rabies can be prevented through vaccination if post-exposure prophylaxis (PEP) is implemented promptly and effectively. Depending on the nature of potential rabies animal interactions, rabies exposure is roughly divided into three categories: I, II, and III. Any exposure deemed to be at risk for rabies requires PEP, including vaccination, thorough cleaning and disinfection of all bites and scratches, local wound infiltration with RIG (only applicable for category III), and timely local treatment of all bites and scratches. The primary purpose of PEP is to prevent the onset of clinical rabies after exposure. PEP regimens typically involve administering human rabies immune globulin (HRIG) once after exposure, followed by the first vaccination on day 0, and subsequent vaccinations on days 3, 7, and 14, ensuring adequate levels of virus-neutralizing antibodies (VNAs) and providing protection against rabies.For immunocompromised individuals, it is recommended to administer a fifth dose on day 28 and to test for seroconversion 7 to 14 days after completing the PEP regimen. For patients who have previously received pre-exposure or post-exposure rabies prevention, only two booster doses of rabies vaccine should be administered on days 0 and 3. Previously vaccinated individuals should not receive HRIG. According to WHO standards, a VNA titer exceeding 0.5 international units per milliliter of serum is considered sufficient to provide protection for humans and animals.
13.2. Rabies Treatment Based on Rabies Immune Globulin (RIG)
Rabies immune globulin (RIG) is a polyclonal or monoclonal antibody specifically targeting G protein epitopes, found to effectively neutralize RABV before it enters the central nervous system (CNS). This provides immediate antibodies until the body responds to the vaccine by actively producing antibodies. RIG comes in two varieties, human rabies immune globulin (HRIG) and equine rabies immune globulin (ERIG), generally administered at doses of 20 IU/kg body weight for HRIG and 40 IU/kg for ERIG to neutralize rabies virus. In general, the area around the wound should be injected with the globulin, ideally on the day of exposure or within 7 days of the first vaccine dose, which helps initially neutralize the virus at the neuromuscular junction.RIG remains a vital component of post-exposure prophylaxis (PEP), as it provides passive immunity during the critical phase before the host generates active immunity. Unfortunately, once the virus enters the central nervous system, these antibodies are limited to crossing the immune-privileged blood-brain barrier (BBB) to combat viral infection. However, studies have demonstrated that simultaneous intravenous administration of RIG and BBB permeable enhancers, such as the cytokine MCP-1 or hypertonic solutions like hypertonic arabinose, effectively allow antibodies to cross the BBB and enter the CNS, neutralizing RABV from the CNS and preventing the development of rabies in mice and rats. Therefore, this therapeutic approach using RIG may represent a promising option for treating rabies..
13.3. siRNA-Based Rabies Therapy
A class of approximately 21-23bp double-stranded RNA molecules, known as siRNA, also referred to as short interfering RNA or silencing RNA, inhibits the expression of specific genes by degrading post-transcriptional mRNA through the RNAi pathway. siRNA-based gene silencing has become a choice for antiviral defenses against many diseases and disorders. Research has focused on plasmids encoding siRNA or siRNA systems based on viral vectors targeting specific RABV genes, which have been constructed and tested for anti-rabies efficacy. In particular, viral vector-based siRNAs typically provide significant protection against lethal RABV attacks and effectively suppress the expression of related genes and RABV proliferation. Recent studies have also highlighted that siRNAs could potentially serve as promising RIG alternatives for rabies treatment.14
Future Prospects of Rabies Vaccines
Recent advances in rabies vaccine development have produced favorable tools for preventing and eradicating rabies. However, in the near future, we will focus on certain areas. The current development of newly invented vaccines is quite considerable; however, developing new vaccine delivery systems remains an important research area that will aid in formulating effective strategic measures to eradicate this disease. Due to the insufficient efficacy of rabies vaccines caused by exposure to adverse temperatures during storage and transportation, the failure of rabies vaccines necessitates research to develop potential rabies vaccines that can be stored and transported outside the conventional cold chain, which will undoubtedly revolutionize vaccine distribution by improving the effectiveness, efficiency, and affordability of vaccine delivery. Recent studies on the thermal stability (SMT) technology of viral vector vaccines will become a new platform for eliminating the cold chain storage requirements of vaccines in the future. Further exploration of novel adjuvants (cPG, colloidal manganese salts, BAFF, etc.) will certainly be promising options for enhancing the efficacy of inactivated anti-rabies vaccines. The application opportunities for creating polyvalent vaccines are promising. Successfully developing bivalent viral vector vaccines for rabies and canine distemper could potentially control both diseases with a single candidate vaccine, especially in developing countries. Furthermore, future vaccine development should aim to create a polyvalent vaccine with broad-spectrum protective efficacy while considering other members of the rabies virus, which will be very effective in reducing this deadliest disease. Despite the gradual improvement in the quality and availability of rabies vaccines and RIGs produced from cell cultures, there has been moderate interest in creating antiviral therapies, ensuring research into potential therapeutic interventions for clinical applications through potential antiviral drugs. Further studies on rabies vaccines using small interfering RNAs (siRNA) in natural hosts or the development of rabies virus therapies based on bispecific antibodies (BsAb) represent more promising options in the treatment of rabies. Additionally, improving post-exposure vaccination and redefining the vaccination regimens of various vaccines remain urgent priorities. Further developing new and improved immunogenic oral bait vaccines in target hosts will be very beneficial for controlling rabies, especially in developing countries through large-scale canine vaccination programs targeting free-roaming dog populations.15
Conclusion
The legendary development of anti-rabies vaccines and the path traversed from Pasteur to the modern immunization era has experienced many ups and downs. Nevertheless, these pioneering efforts have laid a solid foundation for the successful development of vaccines that prevent human deaths and curb canine rabies, thus holding great value. Moreover, utilizing advanced scientific techniques to manipulate the rabies virus genome and the roadmap for developing new vaccine carriers will undoubtedly provide outstanding developments for vaccine research in the near future. However, an urgent priority in rabies-endemic countries is to implement large-scale canine vaccination using injectable vaccines for domestic dogs and easily accessible community dogs, supplemented by oral vaccines for difficult-to-catch free-roaming dogs. This integrated approach can make the global strategic plan to eliminate human rabies deaths caused by dogs by 2030 a reality.
References:Natesan K, Isloor S, Vinayagamurthy B, Ramakrishnaiah S, Doddamane R, Fooks AR. Developments in Rabies Vaccines: The Path Traversed from Pasteur to the Modern Era of Immunization. Vaccines (Basel). 2023 Mar 29;11(4):756. doi: 10.3390/vaccines11040756. PMID: 37112668; PMCID: PMC10147034.
Scan the WeChat QR code to add Pharmacy Time and Space editorPlease indicate: Name + Research Direction!
