
Special Review

Journal:Experimental & Molecular Medicine
Title:Organoids in image-based phenotypic chemical screens
DOI:10.1038/s12276-021-00641-8
Historically, primary hepatocytes are difficult to expand or maintain in vitro. This article reviews the latest advancements in the establishment of hepatocyte organoids and their potential applications in regenerative medicine. First, we provide a background on the renewal of hepatocytes in dynamic balance and damaged livers. Next, we describe insights based on signaling pathways that regulate in vivo hepatocyte renewal, and strategies to establish organoids from primary hepatocytes derived from adult or fetal livers. The characteristics of these organoids will be described. Notably, hepatocyte organoids can be in a proliferative state or a metabolic state depending on the culture conditions. Furthermore, the metabolic gene expression profiles can be regulated according to the principles governing liver zonation. Finally, we discuss the applicability of cell replacement therapy for different types of liver diseases and the current status of hepatocyte transplantation in mouse models. Additionally, we provide insights into how the regenerative microenvironment of the damaged host liver facilitates the repopulation of donor hepatocytes. In summary, in vitro expanded hepatocyte transplantation holds significant clinical application potential in the treatment of liver diseases.
Introduction
The in vitro expansion and maintenance of primary hepatocytes while maintaining their functional characteristics has been a long-standing challenge in the field. Historically, hepatocytes were cultured in monolayers on collagen gels, but using this method, hepatocytes quickly lose morphology and expression of mature hepatic metabolic genes, such as cytochrome P450 (CYPS). Various methods have been adopted to enhance the viability and function of hepatocytes in vitro, including 2D cultures coated with Matrigel or collagen gels (referred to as “sandwich culture”), and co-culturing with non-parenchymal liver cell types (such as Kupffer cells, hepatic stellate cells [HSCs], or liver sinusoidal endothelial cells). The long-term expansion and maintenance of functional primary hepatocytes remain challenging.
Due to the difficulties in culturing primary hepatocytes, different cell sources have been explored, such as directed differentiation of embryonic stem cells (ESCs) or induced pluripotent stem cells (iPSCs) into hepatocytes. In this context, specifying the hepatocyte lineage can be achieved by utilizing factors that activate (or inhibit) specific signaling pathways at different time points during the culture process to mimic the in vivo developmental stages. This approach can generate large quantities of hepatocyte-like cells. However, the resulting cells often lack functional maturity compared to their in vivo counterparts.
In addition to ESC/iPSC-derived cultures, significant progress has been made in establishing organoids from mature cell types derived from adult/fetal progenitors or healthy native tissues. In 2009, the Clevers group described long-term cultures of “mini-intestines” derived from LGR5+ crypt base columnar cells (CbCs) enriched in leucine-rich repeat-containing G protein-coupled receptor 5. LGR5+ CbCs embedded in a 3D extracellular matrix can self-renew and generate various types of intestinal cells under the influence of growth factors such as R-Spin (RSPO), epidermal growth factor (EGF), and Noggin (NOG), spontaneously organizing into crypt-villus structures. Since then, organoid technology has enabled various epithelial cell types to self-renew in vitro, allowing these epithelial cells to expand indefinitely while spontaneously organizing into 3D structures resembling their natural counterparts. Huch and his colleagues described liver organoid cultures derived from intrahepatic cholangiocytes (the epithelial cells forming bile ducts). Since then, other research groups have demonstrated that organoids can be extracted from the common bile duct and gallbladder. However, 3D cultures of primary hepatocytes remain elusive, as primary hepatocytes are the major epithelial cell type performing most hepatic metabolic functions. Importantly, the ability to expand primary hepatocytes in vitro is a key step that enables cell transplantation to treat liver diseases. In a recent breakthrough, Nusse and Clevers’ group described long-term organoid cultures of primary hepatocytes from mice and humans. In this article, we will focus on the strategies for deriving organoids from primary hepatocytes, insights gained from regulating liver homeostasis and regeneration signaling, analyze the characteristics of these organoids from a physiological perspective, and finally introduce the applications of hepatocyte organoids in cell transplantation.
Liver homeostasis and repair: insights into culturing hepatocytes in vitro肝脏动态平衡与修复:体外培养肝细胞的启示
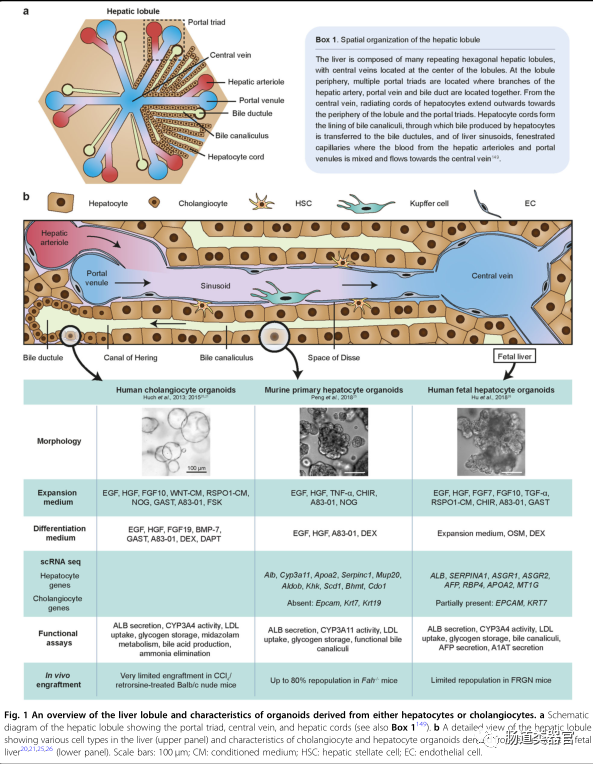
The liver is essential for processes such as metabolism, drug detoxification, and plasma protein production. The two main parenchymal cell types in the liver are hepatocytes and biliary epithelial cells (BECs; cholangiocytes). Hepatocytes perform most liver functions, while bile ducts formed by cholangiocytes transport bile acids produced by hepatocytes to the gallbladder. For an overview of liver structure, see Figure 1. The liver is largely a quiescent organ, with hepatocytes having a long lifespan, estimated to be 200-400 days in rodents. In addition to diploid hepatocytes, the liver also contains binucleate hepatocytes and polyploid hepatocytes, which appear during postnatal development and increase with age, associated with reduced proliferative capacity. Due to their low physiological turnover rate, the liver does not utilize specialized stem cell niches, similar to intestinal crypts, to generate new hepatocytes. Instead, new hepatocytes are generated from the existing pool of mature hepatocytes. However, it remains unclear whether all hepatocytes have self-renewal capability, or whether specific subpopulations of hepatocytes are endowed with unique proliferative abilities. The key question remains: What are the niche signals that regulate hepatocyte turnover and what are the defining characteristics of these hepatocytes?
Recent lineage-tracing studies in mice have provided new perspectives on the turnover of hepatocytes. Hepatocytes adjacent to the central vein (CV) have been observed to preferentially self-renew and replace other hepatocytes in the lobule under the influence of WNT signals (WNT9B and RSPO3) secreted by CV endothelial cells. Hepatocytes surrounding the CV express WNT target gene axis inhibition protein 2 (Axin2) and progenitor cell marker T-box transcription factor 3 (TBX3), which are normally expressed during fetal liver development. These peri-central hepatocytes are predominantly diploid but can also produce polyploid progeny. Over the course of a year of tracking, 20-30% of the lobule was replaced by progeny cells produced by peri-central hepatocytes. While WNT signaling is primarily localized to peri-central hepatocytes in homeostatic livers, activation of the pathway has also been observed in mid-lobular hepatocytes surrounding damaged areas after carbon tetrachloride (CCl4)-induced liver injury. Given the role of the Wnt pathway in liver development and regeneration, as well as its involvement in promoting stem cell activity in various tissues, it is perhaps not surprising that Wnt signaling is associated with hepatocyte proliferation.
In another study, Lin et al. identified a subpopulation of hepatocytes with high expression of telomerase that is widely distributed across the lobule. In this study, cells with high telomerase reverse transcriptase (TERT) produced small clones (up to 8 cells) in healthy mouse livers after 6 months of lineage tracing. An interesting aspect of high TERT cells is their reduced metabolic activity, suggesting that they may have adapted to proliferate rather than perform normal hepatocyte functions. Interestingly, TERT promoter mutations are the most common genetic alteration in hepatocellular carcinoma, with an overall frequency of 60%. Notably, several recent studies using various lineage tracing models have opposed the view that hepatocyte renewal is preferentially localized to the central or portal regions, suggesting instead that it is widely distributed across the lobule. It is also conceivable that multiple and possibly redundant hepatocyte populations can self-renew, as the liver is susceptible to different types of injuries that have differential effects on cells in various parts of the lobule.
A recent study utilizing cellular 14C dating technology on human tissues found that human hepatocytes continuously self-renew throughout adulthood to maintain a relatively youthful liver. Both diploid and polyploid hepatocytes have self-renewal capabilities, but diploid cells have a greater advantage in long-term proliferation compared to polyploid cells. Compared to standard laboratory conditions for feeding mice, this study on human liver tissue may better reflect physiological environments (e.g., frequent exposure to chemicals, pathogens, and drug-induced injuries). Nevertheless, the aforementioned studies in mouse and human tissues provide new evidence that hepatocytes undergo homeostatic renewal at a faster rate than previously thought, with hepatocyte proliferation associated with low ploidy and low metabolic state, and WNT signaling may be the pathway mediating this homeostatic renewal.
Liver regeneration
Compared to the low turnover rate in the quiescent state, the liver has a unique regenerative capacity that is crucial for maintaining liver mass after physical or chemical injury. Events occurring after partial hepatectomy (PHX) in rodents have been well described and serve as a basis for understanding the general principles of liver regeneration. Notably, regeneration after PHX does not rely on a small number of stem cells. Instead, most hepatocytes are capable of proliferating.
Regenerative signals
Following PHX, plasma levels of EGF, insulin, and glucagon (secreted by the intestine, spleen, and pancreas) rise immediately. Hepatocyte growth factor (HGF) precursor peptides are released from the extracellular matrix and activated by urokinase-type plasminogen activator (UPA). These molecules collectively act as powerful mitogens that promote DNA synthesis. Wnt/β-catenin signaling is another important regulatory factor in liver regeneration, promoting G1 phase cell proliferation through cyclinD1 and mitosis. According to our current understanding of WNT pathway regulation, the LGR5/RSPO/zinc and ring finger 3 (ZNRF3) module is likely essential for amplifying WNT signals. Indeed, a recent study confirmed this, showing that liver regeneration is impaired in LGR4/5-deficient mice after PHX. LSECs may be a source of WNT and RSPO. Additionally, inflammatory cytokines are important early signals that initiate liver regeneration. Tumor necrosis factor α (TNF-α) and interleukin 6 (IL-6) are two major cytokines that are typically involved in tissue repair processes across many tissue types. Tumor necrosis factor-α is produced by liver and splenic macrophages in response to lipopolysaccharide, while IL-6 is secreted by liver macrophages and hepatocytes as a result of TNF-α activation. Other signals related to liver regeneration include fibroblast growth factor 7 (FGF-7) and 15 (FGF-15; FGF-19 in humans) and bone morphogenetic protein 7 (BMP-7). There may be redundancy between different signaling pathways, and not all components have been identified.
Intracellular events
In response to regenerative stimuli, various signaling pathways in hepatocytes are activated, leading to cell cycle progression, DNA synthesis, and ultimately cell division. Intracellularly, β-catenin and the intracellular domain of Notch (NICD) translocate to the hepatocyte nucleus within minutes after PHX. Cytokine-induced transcription factors, such as signal transducer and activator of transcription 3 (STAT-3), activated B cell nuclear factor κ light chain enhancer (NF-κB), and activator protein 1 (AP-1) are activated after PHX. YAP1, a key organ size regulator, is also involved in nuclear localization. Although there is no loss of adjacent cells during PHX, changes in shear stress caused by increased blood flow activate YAP1 through the Hippo pathway. Hepatocytes secrete various autocrine and paracrine factors, such as vascular endothelial growth factor, angiopoietin 1 and 2, platelet-derived growth factor, fibroblast growth factors 1 and 2, transforming growth factor α (TGF-α), and granulocyte-macrophage colony-stimulating factor (GMCSF), to promote angiogenesis and stimulate the growth of other liver cell types (hepatic stellate cells, Kupffer cells, and cholangiocytes). Metabolic changes are also a result of regeneration. The expression of genes such as alpha-fetoprotein (AFP), hexokinase, and isoforms of fetal aldolase and pyruvate kinase, which are usually absent in healthy livers, leads to a fetal-like phenotype in regenerating livers.
Termination signals
When the normal liver mass is restored, regeneration gradually terminates. Members of the transforming growth factor-β (TGF-β) family secreted by Kupffer cells and hepatic stellate cells act as brakes on hepatocyte proliferation. Another important inhibitory signal is the suppressor of cytokine signaling (SOCS), which is a negative regulator of various cytokines. In particular, feedback inhibitors SOCS-1 and SOCS-3 of the interleukin (IL)/Janus kinase (JAK)/STAT-3 pathway can inactivate STAT-3, thus terminating hepatocyte proliferation. Other cytokines, such as interferon-γ (IFN-γ), have also been shown to downregulate hepatocyte proliferation. Overall, years of research on liver regeneration have provided important insights into the signals that activate and terminate hepatocyte proliferation. How hepatocytes reconstruct the metabolic gene expression profile of homeostatic liver after regeneration termination remains unclear.
Regenerative signals for expanding hepatocytes in vitro
The combined action of various agents such as EGF, FGF10, HGF, WNT agonists (WNT3a or CHIR99021), Rspo1, gastrin (GAST), NOG, prostaglandin E2 (PGE2), dexamethasone (DEX), Dickkopf-related protein 1 (DKK-1), Forskolin (FSK), and TGF-1 has successfully isolated cholangiocyte organoids from intrahepatic and extrahepatic bile duct cells and gallbladder. A relevant cocktail using EGF, CHIR99021, A83-01, and Y27632 can expand mouse primary hepatocytes in 2D monolayers. Using the cholangiocyte organoid culture medium defined by Huch et al., liver cancer tumor organoid cultures have also been established. However, to date, such media have not yet established primary hepatocyte organoids, prompting us to hypothesize that the media may lack key signals necessary to achieve this goal.
Through studies on in vivo liver regeneration, we speculate that inflammatory signals may be key factors to initiate hepatocyte proliferation in vitro. Indeed, culture media containing tumor necrosis factor-α, along with epidermal growth factor, hepatocyte growth factor, CHIR99021, A83-01, and Y27632, can robustly proliferate mouse primary hepatocytes. Using this scheme, approximately 15% of cultured hepatocytes can form organoids. In the first week of culture, expanded hepatocytes form small spheroids that gradually enlarge to several hundred micrometers over the first three weeks, forming rose-ring-like structures. Notably, they are morphologically distinct from the typical “spherical” structures described in cholangiocyte organoids by Huch et al. and others. Although mature hepatocytes are predominantly binucleate or polyploid, we observed that most of the in vitro expanded hepatocytes are mononuclear, with a high nuclear-cytoplasmic ratio, reminiscent of hepatocytes that regenerate after injury. Hepatocyte-specific marker hepatocyte nuclear factor 4α (hnf-4α) is widely expressed, and bile duct structures can be detected, reminiscent of the structural polarity typically observed in liver lobules. Functionally, hepatocyte-like cells retain key liver functions such as albumin (ALB) secretion, low-density lipoprotein (LDL) uptake, glycogen storage, and CYP3A11 enzyme activity and contain functional bile duct structures. Furthermore, under appropriate culture conditions, hepatocyte-like organoids can upregulate the expression of various metabolic genes. Notably, single-cell RNA sequencing (ScRNAseq) shows that ALB and Cyp3a11 can be induced to levels comparable to mouse primary hepatocytes, indicating that even after long-term culture, they maintain differentiation capacity. In vivo, hepatocyte organoids can be efficiently implanted into damaged mouse livers and reconstruct the expression profile of zonation markers. Finally, hepatocyte organoids can be continuously passaged and expanded for at least 8 months (the longest culture involved ultra-low temperature preservation after about a year of culture, unpublished data). To our knowledge, this is the first study demonstrating long-term 3D culture of hepatocyte organoids that retains key functional characteristics and in vivo regenerative capabilities.
Cytokine dependency and mode of action
Interestingly, we found that tumor necrosis factor-α is a key factor in establishing hepatocyte organoid cultures. Substituting tumor necrosis factor-α with another inflammatory cytokine IL-6 did not produce any organoids. Moreover, the long-term expansion of hepatocytes remains dependent on tumor necrosis factor-α. At any time point in culture, removal of tumor necrosis factor-α leads to restricted hepatocyte growth, ultimately resulting in culture deterioration. In vivo, tumor necrosis factor-α is generally considered an important signal for liver regeneration. For example, antibody blockade of tumor necrosis factor-α inhibits regeneration, while tumor necrosis factor receptor 1 knockout mice exhibit delayed regeneration. Thus, the critical role of tumor necrosis factor-α in maintaining in vitro cultures is not surprising.
Early hepatocyte culture studies provided some insights into the role of tumor necrosis factor-α in hepatocyte proliferation. One study found that while epidermal growth factor alone could not promote the proliferation of cultured hepatocytes, the combined use of epidermal growth factor and tumor necrosis factor-α could stimulate approximately 35% of cultured hepatocytes to enter the S phase of the cell cycle through cyclin D1 and cyclin-dependent kinases 1 and 2. Furthermore, tumor necrosis factor-α inhibited the maturation of fetal liver cells induced by the suppressor M, promoting the cell cycle progression of cultured cells. It is known that tumor necrosis factor-α can also prevent apoptosis through the NF-κB signaling pathway, promoting cell survival and proliferation. Indeed, inhibition of NF-κB leads to culture deterioration, suggesting that TNF-α signaling is likely mediated by the NF-κB pathway. Importantly, there is significant crosstalk between the NF-κB and Wnt signaling pathways.
A recent study by the Dier laboratory indicated that in patients with severe alcoholic hepatitis and liver injury mouse models, inflammatory cytokines (such as tumor necrosis factor-α) can reprogram mature hepatocytes into proliferative hepatocytes with fetal-like metabolic characteristics. The acquisition of a fetal profile is mediated by the inhibition of the epithelial splicing regulatory protein 2 (ESRP2)-mediated adult splicing program, leading to the accumulation of fetal splicing variants of genes such as NF2 and CSNK1D, which reduce the kinase activity of YAP1 degradation. Consequently, the expression of YAP1 target genes AREG, CTGF, and PTGS2 is upregulated. Consistent with in vivo observations, qrt-pcr and splicing variant analyses showed that hepatocyte organoids cultured in TNF-α media significantly downregulated Esrp2, accumulating fetal RNA splicing variants (Nf2, SLK, Fln, and Kras) and upregulating the aforementioned YAP1 target genes. Importantly, after withdrawal of tumor necrosis factor-α, hepatocyte organoids reconstructed the expression of Esrp2 and upregulated adult liver-specific markers such as ammonia detoxification pathways and coagulation factors. In summary, this study provides a potential mechanism by which tumor necrosis factor-α reprograms adult hepatocytes into fetal-like hepatocytes.
Cytokine-induced in vitro regeneration beyond the liver
The liver is not the only organ that relies on inflammatory signals for regeneration after injury. In fact, in multiple tissues, such as skeletal muscle, intestine, colon, hair follicles, skin, and the central nervous system, inflammation and regeneration are closely coupled. Inflammatory cytokines TNF-α and interleukins trigger regenerative responses in adult progenitor or mature cells by activating various transcription factors (such as NF-κB, JAK/STAT, AP-1, YAP1, and NOTCH), thereby activating transcription programs that promote cell survival, proliferation, dedifferentiation, and acquisition of fetal-like phenotypes. These observations lead us to hypothesize that regenerative cytokines may have a more universal role in cell cycle regulation and could potentially be utilized beyond injury contexts to expand otherwise quiescent/slow-cycling primary cell types in vitro. Indeed, cytokines have also been used to extend the regeneration effects in vitro for other tissue types, namely muscle and lung, both of which are known to experience cytokine-driven regeneration after injury. In fact, muscle stem cells (satellite cells) can long-term expand under the combined action of various cytokines (TNF-α, IL-1α, IL-13, and IFN-γ), while alveolar type II epithelial cells proliferate vigorously in the presence of IL-1 and TNF-α. Overall, these studies provide new evidence supporting the role of cytokines in the in vitro culture of other “hard-to-culture” cell types.
An alternative strategy: fetal-derived human hepatocytes
In a parallel study by the Clevers group, Hu et al. used a cocktail including EGF, HGF, Fgf7 and 10, WNT agonists (CHIR99021 and Rspo1), A83-01, and Y27632 to expand mouse hepatocytes. FGF7 is a niche factor previously found to be important for liver regeneration. Interestingly, Hu et al. observed that cells surrounding the CV had slightly higher organoid formation efficiency than other hepatocytes in the lobule. Diploid and tetraploid hepatocytes can form organoids, while octaploid cells cannot. Morphologically, hepatocyte organoids resemble “grape-like” cell clusters rather than the denser packing structures observed in hepatocyte organoids by Peng et al. Importantly, hepatocyte organoids can perform major functions such as albumin secretion, CYP1A2 activity, glycogen storage, and low-density lipoprotein uptake, showing extensive bile duct networks. The culture medium supports organoid formation, but only a small fraction of cells form hepatocyte organoids (0.5-1%). The rate of organoid formation in this medium is significantly lower than that in media containing TNF-α. Notably, organoids can be preserved for up to 3 months, after which they show limited growth.
To culture human hepatocytes, Hu et al. adopted a different strategy, using fetal-derived hepatocytes, which are expected to have higher proliferative capacity than mature hepatocytes, making them more suitable for long-term expansion. Previous studies have shown that fetal hepatocytes can maintain mature liver functions while being cultured for months. In the current study, fetal hepatocytes were obtained from donor fetuses at 11-20 weeks of gestation using a two-step perfusion method, which typically enriches hepatocytes in adult livers. Approximately 1% of plated cells can form organoids. In this study, no identity of the cells producing organoids (or their corresponding developmental stage) was determined. Using a culture medium similar to that for mouse organoids, human fetal liver-derived hepatocyte organoids can expand for nearly a year. Transmission electron microscopy images show the aggregation of glycogen granules, displaying cellular characteristics (large nuclei, prominent nucleoli, numerous mitochondria, few pools, bile ducts, tight junctions, etc.). This is very similar to hepatocytes, indicating their maturation in vitro.
Adult human hepatocyte culture remains challenging
In the same study, Hu et al. established hepatocyte organoids from human hepatocytes derived from adults and children. However, compared to fetal-derived hepatocytes or adult mouse primary hepatocytes, the proliferative capacity of mature hepatocytes seems limited (2-2.5 months). Since our work was published, several groups have reported monolayer cultures of 95-100 human hepatocytes. Zhang et al. found that the expression levels of aging and cell cycle arrest genes significantly increased during culture, with adult hepatocyte growth being impeded after 4 generations (equivalent to approximately 300-fold expansion). To counteract this cellular aging effect, cells were cultured under low-oxygen conditions, resulting in a 10,000-fold expansion (up to 8 generations). Additionally, Katsuda et al. attempted reprogramming methods based on previously described small molecules to induce proliferation in adult hepatocytes. They found that the proliferation of adult hepatocytes was very limited. Subsequently, Katsuda et al. focused on infant hepatocytes, which have been shown to be easier to expand in culture. After treatment with maturation-inducing factors OSM and DEX, these cells can differentiate into cells expressing various functional CYP enzymes (such as CYP1A2, CYP2B6, and CYP3A4) at levels comparable to mature hepatocytes. Furthermore, as the authors noted, although the expression and functional activity of liver markers (such as CYP enzyme activity) gradually decline during culture, they can be passed on for more than 10 generations. These studies and several recent others highlight the challenges of long-term expansion of adult hepatocytes.
Cell identity and state: bipotent or not?
The characteristics of hepatocytes cultured in vitro have been an interesting question, as early studies found that hepatocytes can adopt biliary fates after injury and in vitro culture. Michalopoulos et al. were the first to provide evidence of in vivo hepatocyte conversion to cholangiocytes, demonstrating that dipeptidyl peptidase IV (DPPIV)+ transplanted hepatocytes acquired cholangiocyte morphology after inducing DPPIV- host rat cholangiocyte injury. In a later study, Yanger et al. used lineage tracing in a mouse model of cholestatic liver injury to demonstrate that hepatocytes can transdifferentiate into duct-like cells during cholangiocyte injury. Additionally, Schaub et al. demonstrated that in a mouse model of Alagille syndrome (a disease characterized by impaired formation of peripheral bile ducts), partially functional biliary trees still formed. Researchers used lineage tracing to show that these ducts were of hepatocyte origin, and in this disease model, hepatocytes were able to form new ducts. In summary, these studies suggest that when cholangiocyte proliferation is inhibited, hepatocytes have the ability to transition to a ductal state.
Single-cell RNA-SEQ can be used to evaluate the heterogeneity among cultured hepatocytes. According to scRNA-seq, Hu et al. identified five different cell clusters in their mouse hepatocyte cultures, defining them as non-cycling mature hepatocytes (Albhigh), cycling progenitor cells with high levels of precursor markers (secreted phosphoprotein 1 [Spp1] high, Allow), more primitive progenitor cells with high levels of cycling genes but lacking mature markers (two clusters), and some cells with bile duct markers (keratin 7 [Krt7]+). Consistent with the transcriptional data, spontaneously formed cystic organoids resembling biliary organoids were observed in cultured ALB+ labeled cells. Furthermore, hepatocytes cultured in bile duct media strongly upregulated prototypical biliary markers (Krt7 and Krt19), while downregulating hepatocyte markers (hnf-4α and alb).
The conversion of hepatocytes to bile duct cells is consistent with previous studies in monolayer cultures. Katsuda et al. demonstrated that mature hepatocytes can be reprogrammed into bipotent progenitor cells using small molecules (CHIR99021, Y27632, and A83-01) and can be induced to upregulate either hepatocyte or biliary markers. Under culture conditions promoting biliary fate, biliary markers such as KRT19, aquaporins (AQPs) 1 and 9, as well as ion channel proteins cystic fibrosis transmembrane conductance regulator (CFTR) and anion exchanger 2 (AE2) are strongly induced. The resulting duct-like structures secrete fluorescent dyes into the lumen and can respond to hormone secretin by expanding the luminal space, indicating that BEC function is normal.
Additionally, Peng et al.’s scRNA-seq showed that hepatocyte-specific markers (e.g., hnf-4α, Alb, apolipoprotein A1, transthyretin, and Serpina1c) are widely expressed across the cultured hepatocyte population. The cultured cells are enriched with genes associated with cell lysis, lipoprotein particle remodeling, coagulation, retinol metabolism processes, G1/S phase transition regulation, and responses to tumor necrosis factor. Additionally, some cells highly express genes related to mitosis, cell cycle regulation, chromosome segregation, remodeling, and DNA replication, indicating that hepatocytes are cycling. A major distinction from other cultured cells is the lack of typical biliary markers (such as epithelial adhesion molecules, Krt7, Krt19, Aqp1, Aqp4, and claudin7), suggesting that the vast majority of the cells cultured with TNF-α are hepatocytes without signs of transdifferentiation. In summary, the aforementioned studies suggest that cultured hepatocytes possess cellular plasticity, and the addition of appropriate signals can suppress biliary fates, producing only hepatocytes.
‘Proliferative’ vs ‘metabolic’ states
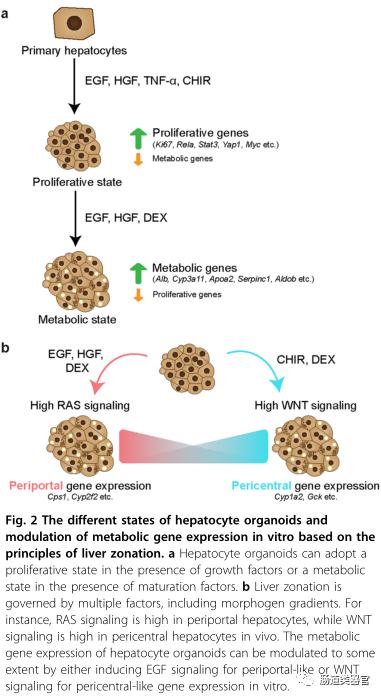
An interesting aspect of hepatocyte organoids is that in vitro culture encapsulates certain aspects of liver regeneration. RNA-SEQ data supports this, as it compares hepatocyte organoids to hepatocytes isolated from livers induced by PHx. We noted that transcription factors typically associated with tissue regeneration, previously described as key factors in liver regeneration post-PHx, were upregulated, such as in vivo. For example, transcription factors RelA (NF-κB p65 subunit), STAT3, YAP1, Myc, Jun, and Fos (forming the AP-1 complex) were detected in organoids, but were at low levels or absent in primary hepatocytes isolated from healthy mouse livers. Acute phase proteins and cytokines associated with inflammatory responses are highly expressed in proliferating hepatocytes in vitro. Additionally, hepatocyte organoids secrete growth factors, including insulin-like growth factor 2 (IGF2), transforming growth factor-α, and angiogenic factor VEGF-A. A major outcome of “in vitro regeneration” is the downregulation of metabolic genes that are typically highly expressed in the liver, indicating an adaptation to a “proliferative” state. For instance, compared to adult mouse hepatocytes, the expression levels of CYP enzymes (Cyp3a11 and Cyp2e1), apolipoproteins (APOA2 and APO3), serine protease inhibitors (Serpin1e and Serpin3k), and major urinary proteins (Mup3 and Mup20) are lower. To test whether proliferative hepatocytes re-express functional genes after cessation of expansion and inflammatory factors, Wnt and TNF-α were removed from the culture media, and dexamethasone was added to promote hepatocyte maturation. Indeed, when hepatocytes cultured in expansion media (2-3 months) switched to differentiation media for 3-5 days, the cultured cells upregulated a set of genes corresponding to mature hepatocyte functions. ScRNA-seq showed that hepatocyte markers Alb (the most abundant plasma protein), Cyp3a11, SERPINA1, Aldob, KHK, Bhmt, and Mup20 were induced to levels comparable to primary hepatocytes. Furthermore, functional assays such as albumin secretion and CYP3A11 enzyme activity were performed on hepatocyte organoids cultured for 3-7 months, confirming scRNA-seq data. Subsequently, markers associated with regeneration and proliferation were downregulated, which correlated with reduced proliferation of organoids in differentiation media.
A recent single-cell atlas study investigated the transcriptional landscape of hepatocytes after PHX, providing evidence that hepatocytes briefly re-activate transcriptional programs similar to those experienced during a strong proliferative phase, with a set of different cells becoming “metabolically hyperactive” to compensate for any temporary deficits in liver function. This division of labor ensures that the liver continues to function optimally while undergoing tissue repair. In summary, these studies suggest that hepatocytes can be in either a “proliferative state” or a “metabolic state” in both vivo and vitro. More importantly, hepatocyte organoid cultures retain the ability to upregulate the expression of various metabolic markers that are typically seen in mature hepatocytes. However, it should be noted that the metabolic profile of the adult liver is highly zonated and regulated by various factors, many of which are absent in the current culture conditions. In the following sections, we will discuss how understanding the pathways regulating liver zonation and ecological niche signals present in the liver microenvironment can modulate the expression profiles of hepatocytes in vitro.
Modulating hepatocyte function in vitro: from the perspective of liver zonation
Hepatocytes within liver sinusoids are exposed to different gradients of morphogens, oxygen, nutrients, hormones, and metabolites, leading to zonated gene expression. Cells surrounding the portal vein (PV) express different gene subsets than those surrounding the central vein (CV). Wnt/β-catenin signaling is generally considered one of the main determinants of spatial zonation of gene expression, known as “zonation maintainers.” The production of WNT9B and RSPO3, as well as WNT2 expression from sinusoidal endothelial cells extending from the CV, leads to a gradient of WNT morphology in the lobule. Conversely, the activity of Wnt/β-catenin is inhibited by negative regulators of the portal region. As shown by Benhamouche et al., blocking WNT signaling can promote gene expression around the portal vein, while knocking out Apc promotes gene expression around the central vein. While WNT ligands are enriched in the peri-central region, RAS signaling is considered one of the major pathways regulating gene expression around the portal vein. It is likely that many other pathways that may contribute to liver zonation remain to be identified.
Although it is desirable to reconstruct liver zonation in vitro to achieve functional diversity among cultured hepatocytes, this has technically been challenging due to the lack of spatially localized ecological niche signals and morphogen gradients typically observed in native tissues. Additionally, due to the insolubility and low expression yield of WNT proteins, they have traditionally been difficult to manipulate. Significant advances have been made by designing more easily manipulatable effective soluble WNT agonists. WNT agonists are designed to bind LRP6 and FZD receptors, for example, by attaching pan-Frizzled antibodies to the CT domain of the WNT antagonist Dkk1. Recently, WNT agonists have been shown to maintain various epithelial organoids in vitro. Specifically, we demonstrated that using next-generation WNT substitutes, according to the previously described scheme, hepatocyte organoids can be well expanded. Additionally, earlier work by Habib et al. showed that immobilized WNT, i.e., WNT tethered to beads, can serve as a stem cell niche to mediate asymmetric division. In a recent study, the Lutolf laboratory demonstrated that micro-patterned scaffolds mimicking crypt-villus structures can be used to maintain mini-intestines in vitro. In summary, micro-patterned scaffolds co-cultured with non-parenchymal cell types, tethering WNT agonists to a “signal hub” mimicking the CV, and modulating morphogen flow may enable hepatocyte culture to achieve spatial organization and gradient gene expression similar to liver lobules.
Hepatocyte transplantation to treat liver diseases
Orthotopic liver transplantation is currently the preferred treatment for end-stage chronic liver disease. Due to a severe shortage of donor organs, hepatocyte transplantation has been explored as an alternative strategy. Cryopreserved primary human hepatocytes have been used for transplantation in various diseases, such as acute liver failure, coagulation factor VII deficiency, and Crigler-Najjar syndrome. However, due to a lack of high-quality donor cells, implantation efficiency is limited, and despite the application of immunosuppressants, transplanted cells still face immune rejection, leading to a loss of clinical benefit over time, hindering the widespread clinical application of hepatocyte transplantation. Here, we discuss the types of liver diseases that cell transplantation may potentially treat, the potential uses of in vitro expanded hepatocytes in cell transplantation, and strategies to improve their long-term engraftment.
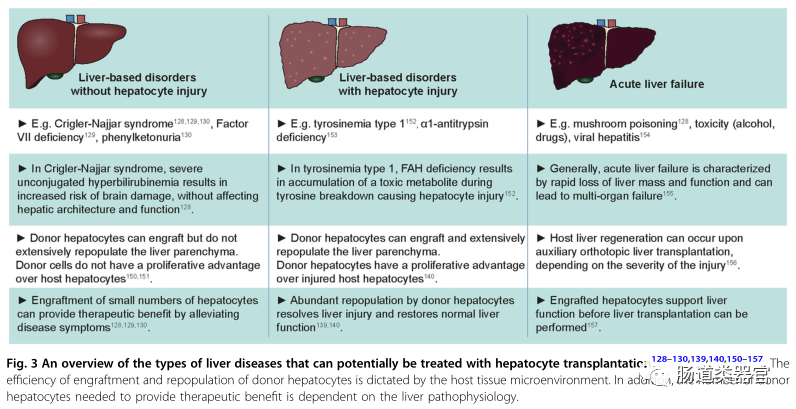
Types of liver diseases that can potentially be treated by hepatocyte transplantation
The efficiency of hepatocyte repopulation in the host liver and the extent of liver repopulation required for therapeutic benefit varies among different types of liver diseases, depending on the pathophysiology of the affected host liver. For example, many liver diseases are caused by single-gene defects, including Crigler-Najjar syndrome (hyperbilirubinemia), coagulation factor VII deficiency (coagulation disorders), and phenylketonuria (accumulation of phenylalanine in the blood). The lack of functional gene products impedes the execution of certain essential liver functions without damaging the liver itself. In such cases, a small number of transplanted hepatocytes may provide therapeutic benefits and alleviate disease symptoms by producing the necessary functional proteins. The number of transplanted donor cells required for clinical benefit in these diseases is not clear, but several patients have shown symptom relief after receiving a number of cells equivalent to 5-10% of liver mass. For instance, a patient with Crigler-Najjar syndrome underwent transplantation of approximately 4% of total liver mass, significantly reducing serum bilirubin levels and maintaining relief for 6 months, after which the patient ultimately received a whole liver transplant. However, over time, the loss of clinical benefits indicates a lack of long-term donor cell engraftment in the host liver.
Cell transplantation with in vitro-expanded hepatocytes
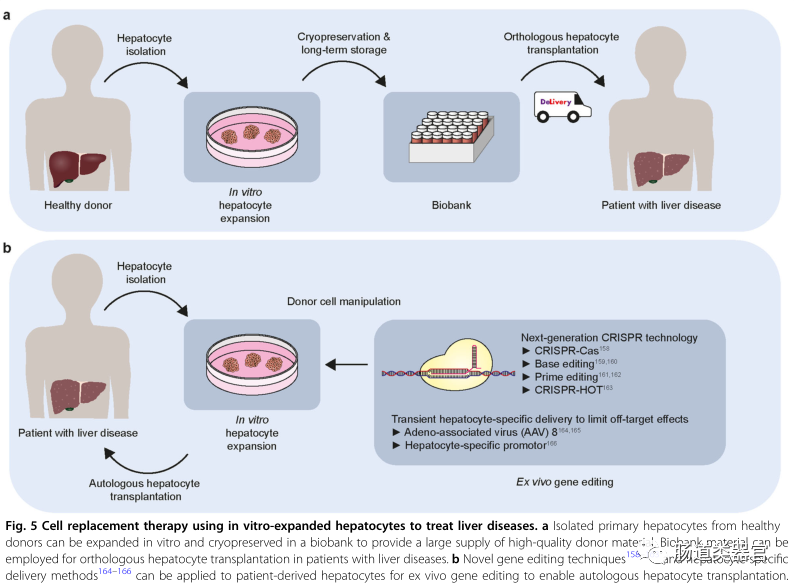
To improve the clinical outcomes of human hepatocyte transplantation, major challenges need to be addressed, such as the lack of suitable, healthy, and high-quality primary cells for transplantation, difficulties in preservation, low implantation and repopulation rates, and a lack of long-term effects on organ function recovery. To date, cryopreserved primary human hepatocytes have been used in clinical applications of hepatocyte transplantation. While cryopreservation facilitates the long-term storage of donor materials, cryopreserved hepatocytes show reduced cell viability, cell attachment efficiency, and albumin production compared to freshly isolated primary hepatocytes, leading to a decline in the quality of cell transplantation. Cultured hepatocytes are more tolerant of low-temperature preservation than freshly isolated primary hepatocytes. Rodent hepatocytes cultured for 24 hours in monolayers before cryopreservation show higher survival rates and culture efficiencies after thawing compared to primary hepatocytes that were directly cryopreserved after isolation. According to our own experience, hepatocyte organoids can withstand repeated freeze-thaw cycles for in vitro expansion. Cultured hepatocytes can be transported overnight in a freezing medium containing 2% fetal bovine serum, maintaining high survival rates (>90% survival, unpublished data) for mouse transplantation.
Engraftment of in vitro-expanded primary hepatocytes in liver injury mouse models
Cholangiocyte-derived organoids
Research by Huch et al. demonstrated that cholangiocyte-derived organoids can be induced to express hepatocyte markers (such as hnf-4α, albumin, and multidrug resistance-associated protein 4) in vitro and exhibit certain liver functions (such as low-density lipoprotein uptake and glycogen storage). However, these “transdifferentiated” cholangiocyte organoids showed limited repopulation in Fah−/− mice (with less than 1% liver parenchyma). Similarly, human cholangiocyte organoids were only implanted in the form of single and double cells and showed no evidence of repopulation in BALB/c nude mice. The low repopulation rate of cholangiocyte organoids in the liver is likely a result of poor transdifferentiation of cholangiocytes to hepatocytes before and/or after implantation. In line with this, in vivo observations have rarely noted cholangiocyte to hepatocyte transdifferentiation during liver regeneration, only observed in extreme cases where hepatocyte proliferation is severely impaired in transgenic mouse models. The lack of implantation and redifferentiation of cholangiocyte-derived organoids underscores the necessity of using primary hepatocyte organoids as the cell type for transplantation.
Primary hepatocyte organoids
Transplanting mouse hepatocyte organoids into Fah−/− mice demonstrated the in vivo regenerative capacity of in vitro expanded primary hepatocytes. The transplanted hepatocytes massively repopulated the damaged host liver after approximately 100 days, resulting in FAH+ clones occupying up to 80% of the liver parenchyma. Donor cells expressed hepatocyte markers hnf-4α but did not express biliary markers sox9, krt7, or kr19. Notably, the hepatocyte organoids were cultured in media promoting expansion before transplantation. Interestingly, donor cells underwent maturation in vivo and reestablished zonation marker expression. For example, cells near the CV expressed markers such as glutamine synthetase (GS), glutamate transporter 1 (GLT1), and CYP2E1, while cells outside the CV region expressed CPS1. Importantly, none of these markers were detected in the host tissue. Overall, these observations suggest that the expression pattern of donor hepatocytes at the time of transplantation is determined by the host liver microenvironment.
Hepatocyte organoids derived from human fetal liver
Hu et al. successfully implanted human fetal liver-derived hepatocyte organoids into Fah−/−/RAG2−/−/IL2rγ−/− (FRG) mice (i.e., non-obese diabetic (NOD) strain FRG-deficient mice), although to a limited extent. Nevertheless, the transplanted cells expressed functional markers such as ALB, CYP2E1, and MRP2, but not fetal markers AFP, indicating maturation in vivo. The limited range of implantation is unclear and may be due to a lack of maturation in vivo. To verify this hypothesis, the authors compared the implantation of fetal-derived organoids with primary hepatocytes and organoids from the same pediatric source, noting that primary hepatocytes and organoids significantly outperformed fetal-derived organoids in implantation and repopulation. This result suggests that mature hepatocytes are more suitable for cell transplantation.
2D culture of primary murine/human hepatocytes
In addition to in vitro expanded primary hepatocyte organoids, earlier research by Katsuda et al. indicated that chemically induced liver progenitor cells (CLIPS) derived from rat hepatocytes maintain high repopulation capacity in vivo. Using cDNAuPA/severe combined immune deficiency (SCID) mice, specifically expressing uPA in hepatocytes under the action of the albumin promoter, inducing severe liver injury, CLIPS can replant up to 90% of the host liver and express hnf-4α, multidrug resistance-associated protein 2 (Mrp2), and various CYP enzymes, indicating a high degree of in vivo differentiation. Building on their previous work, Katsuda et al. demonstrated that human juvenile hepatocytes could be reprogrammed into human segments in culture, showing highly variable repopulation efficiency. In some mice, over 90% of liver parenchyma achieved redifferentiation, and the cells expressed functional markers MDR1, TTR, GS, CYP1A2, and CYP3A4, indicating in vivo maturation. In another study, Zhang et al. demonstrated that cryopreserved human hepatocytes expanded in vitro could largely repopulate the livers of FRG mice. Donor cells expressed hepatocyte markers hNF-4α, ALB, CYP3A4, and GS. Notably, the latter two studies indicated that the repopulation capacity of expanded human hepatocytes declines with prolonged culture time, as the repopulation capacity after transplantation decreases compared to earlier generations. These studies suggest that suboptimal culture conditions may to some extent impair the implantation potential of the cells.
Regenerative host liver microenvironment facilitates donor cell repopulation
How hepatocytes repopulate the host liver remains a fascinating question. Previous studies by the Grompe laboratory and others have shown that transplanted primary hepatocytes can replace up to 90% of host mouse liver in Fah-deficient or FRG-deficient mice, both widely used for liver repopulation studies. Considering that approximately 1 million cells (or 2% of total liver mass) are typically transplanted into mice, and it is estimated that only 10% of injected cells engraft in the liver, significant expansion is required in vivo to reach the observed repopulation levels. In these studies, FAH-−/− mice have been continuously administered NTBC in their drinking water from birth to prevent the production of toxic metabolites and the occurrence of liver damage. After transplantation, NTBC is removed to initiate liver damage. In nFah−/− mice, the regeneration of host hepatocytes is impaired, but donor cells are unaffected, providing a proliferative advantage to donor cells. It is speculated that the regenerative, signal-rich liver microenvironment promotes the proliferation of donor cells.
In Fah−/− mice transplanted with hepatocyte organoids, FAH+ grafts of varying sizes were observed, likely arising from clonal repopulation of transplanted cells. To track Wnt activity in donor cells, hepatocytes carrying Axin2-rtTA and TetOH2B:GFP reporter genes were transplanted into Fah−/− mice. In the two days prior to analysis, transplanted mice were treated with doxycycline to activate Axin2 expression in donor hepatocytes. GFP expression was detectable in FAH+ clones but not in host cells. In smaller FAH+ clones, many GFP+ cells were scattered throughout the clones, indicating responsiveness to WNT signals. In contrast, in larger grafts, GFP+ cells were confined to the donor cells at the graft-host tissue interface, absent in the center of the graft. Consistent with this observation, the expression of Wnt2, Wnt9b, and Rspo3 was largely upregulated in the host tissue. The expression of RSPO3 is typically limited to endothelial cells surrounding the vasculature in uninjured livers but is upregulated in the endothelial cells of host tissue surrounding the clone boundary. It is likely that the Wnt-rich environment in the host liver promotes the proliferation of donor cells, consistent with the role of Wnt signaling in other regenerative paradigms.
Previous studies have indicated that angiogenesis is a key aspect of liver regeneration and function. VEGF secreted by regenerating hepatocytes promotes LSEC proliferation. Neutralizing VEGF not only impairs endothelial cell proliferation but also indirectly harms hepatocyte proliferation. Furthermore, studies have shown that LSECs establish an inductive vascular niche during liver regeneration after partial hepatectomy by activating VEGF receptor 2 (VEGFR2) and upregulating the transcription factor DNA binding inhibitor 1 (ID1), leading to the production of WNT2 and HGF, stimulating hepatocyte proliferation. These observations are consistent with the crosstalk between hepatocytes and LSECs, which are the main source of WNT-RSPO signaling for hepatocytes. In addition to the crosstalk between endothelial cells and hepatocytes, another example of paracrine signaling between host tissue and donor hepatocytes has been described. After donor hepatocyte transplantation, host hepatocytes upregulate IGF2 to support liver regeneration. In vivo blockade of IGF2 impairs liver regeneration, while its activation leads to increased rates of hepatocyte implantation and the number of MKI67-positive donor cells. The crosstalk network between donor hepatocytes, host hepatocytes, and the host microenvironment likely determines the repopulation of donor hepatocytes.
The contribution of liver stem cells to hepatocyte transplantation has been studied in preclinical transplantation settings, where the animal models used do not allow for selective repopulation of donor cells. After hepatocyte transplantation in DPPIV-deficient rats, an increase in the number of host HSCs was observed post-transplantation, primarily near the transplanted DPPIV donor cells, suggesting close interactions and potential crosstalk between HSCs and donor hepatocytes during donor cell transplantation. In later studies, primary human hepatocytes were co-transplanted with isolated human hematopoietic stem cells or hematopoietic stem cell lines (LX-2) into SCID mice. Transplantation of human hepatocytes alone led to scattered human ALB+ cells implanted throughout the parenchyma, while co-transplantation resulted in increased grafting, which was more pronounced for LX-2 cells and activated HSCs compared to quiescent HSCs. In summary, these studies indicate that close interactions between hepatocytes and hematopoietic stem cells participate in regulating hepatocyte implantation during transplantation, although the exact mechanisms and signaling pathways involved, as well as whether hematopoietic stem cells also assist donor cells in supporting liver reconstruction, remain to be elucidated. While the role of HSCs in liver regeneration is still unclear, it is known that HSCs respond to paracrine signals generated by hepatocyte regeneration and other cell types (Kupffer cells and ISECs). In this process, the importance of HSCs in liver regeneration following partial hepatectomy has been noted. It is possible that similar crosstalk between hematopoietic stem cells and donor hepatocytes also contributes to donor cell implantation and reconstruction.
Kupffer cells and other immune cells play a complex role in hepatocyte transplantation, as they primarily participate in the clearance of transplanted cells and the prevention of transplantation, which is beyond the scope of this article and has been discussed elsewhere. On the other hand, Kupffer cells are a major source of TNF-α and IL-6, two inflammatory cytokines involved in liver regeneration, supporting hepatocyte proliferation in vitro. Therefore, the supportive role of Kupffer cells in the process of in vivo transplanted hepatocyte regeneration is also to be expected.
Conclusions
The ability to culture or expand appropriate types of cells to provide a large number of healthy donor cells is the first necessary step to make cell replacement therapy feasible. Recent breakthroughs in expanding in vitro primary hepatocytes while retaining their in vivo regenerative capacity represent an important milestone toward achieving hepatocyte transplantation. Furthermore, the ability to expand, freeze, and transport hepatocytes without compromising their quality will enable the creation of a biobank that ensures the availability of hepatocyte transplantation (in vitro expanded) donor materials, thus addressing the donor shortage issue. More importantly, autologous cell transplantation will become feasible as new gene editing technologies and liver cell-specific delivery methods can be applied to patient-derived hepatocytes for in vitro gene editing to restore disease-specific mutations. While currently, the lack of sufficient donor cells implanted in the host liver or the lack of long-term maintenance may pose significant challenges to the long-term efficacy of cell transplantation, we anticipate that understanding the mechanisms by which donor hepatocytes repopulate diseased host livers will provide in-depth strategies to overcome these limitations. We are optimistic that in the foreseeable future, in vitro expanded primary hepatocytes will be widely applied in cell replacement therapies for liver diseases.
Join 6000+ peers to focus on organoids!
The Intestinal Organoids is committed to publishing high-quality information in this field. If you have particularly liked and appreciated organoid research papers, reviews, or personal opinions, or if your research team has published new articles and technological advances in the field of organoids, or if your team needs talent related to organoid technology. You are welcome to send messages or emails to the public account editor in this subscription number:[email protected][Reply within 12 hours], we open communication and free release and obtain information, and welcome everyone to provide high-quality information, recruitment information, and personal reviews!

#What is the ISCO program?
By joining the “ISCO program”, you will achieve the exchange and sharing of organoid strains at extremely low cost, achieving standardized organoid preparation and detection based on the zlab and intestinal organoids public account platform, ensuring that your research proceeds smoothly and with high quality. We will include you in the ISCO program member WeChat group, where all your organoid questions can be answered and helped in the most timely manner, allowing you to communicate with all members participating in the ISCO program, exchanging organoid strains and stem cell research tool resources.
Professional research teams and independent PIs are welcome to joinISCO program professional team member group, add the initiator WeChat, you can be invited to join the group.
Email:[email protected]
WeChat:zlabxyz
Or scan the QR code below

Ordinary organoid technology enthusiasts, students, and researchers can join our technical exchange group:
